Роль макрофагов и цитокинов в развитии сахарного диабета типа I
Сахарный диабет типа I представляет собой аутоиммунное заболевание, которое вызывается развитием воспалительной реакции непосредственно в поджелудочной железе или около нее. В воспалительных инфильтратах выявляются макрофаги, моноциты, Т-лимфоциты; на начальном этапе индуцированного у экспериментальных животных диабета ткань железы инфильтрирована преимущественно активированными макрофагами.
Истощение пула мононуклеарных фагоцитов предварительным введением неорганических частиц предотвращало развитие диабета у животных [1251]. Диета с низким содержанием ненасыщенных жирных кислот также приводила к снижению содержания макрофагов в поджелудочной железе и препятствовала развитию аллоксан-индуцирован- ного диабета у мышей [1127]. Это указывает на ключевую роль макрофагов в повреждении инсулинсекретирующих β-клеток. Действительно, инкубация активированных перитонеальных макрофагов с клетками поджелудочной железы ингибировала синтез инсулина и в последующем вызывала гибель как α-, так и β-клеток, при этом цитотоксический эффект макрофагов снижался удалением из культуральной среды L-аргинина или добавлением ингибиторов NO-синтазы [938]. В то же время инактивация ИЛ-1 и ФНО-α с помощью специфических антител не влияла на цитотоксичность. На модели индуцированного стрептозотоцином диабета у животных также было показано, что введение ингибитора NO-синтазы L-NMMA предотвращает повреждение клеток поджелудочной железы и развитие диабета [1058]. Эти данные указывают на то, что повреждающее действие макрофагов на клетки поджелудочной железы осуществляется через продукцию NO-радикалов. Действительно, у мышей, трансгенных по NO-синтазе, наблюдается быстрая деградация клеток поджелудочной железы, и уже через 4 недели после рождения развивается гипоинсулиновое состояние, сходное с диабетом типа I [1598].При изучении действия активированных макрофагов на клетки поджелудочной железы ингибирование цитокинов (ИЛ-ф и ФНО) прямо не влияло на цитотоксичность, что можно объяснить высокой продукцией NO-радикалов фагоцитами.
Однако в 1985 г. было впервые показано, что культуральная среда, полученная от активированных человеческих моноцитов, снижает продукцию инсулина и вызывает деструкцию β-клеток в островках Лангерганса, выделенных у крыс и человека [1090]. Наиболее активный компонент среды был идентифицирован как ИЛ-1 [319]. Дальнейшие исследования показали, что инкубация островков Лангерганса крыс с ИЛ-1 в течение 1—3 часов усиливала глюкозостимулированную секрецию инсулина, однако продолжительная экспозиция (в течение 15—18 часов), наоборот, на 80—90 % снижала продукцию инсулина; это ингибирование было обратимым и секреция восстанавливалась через 3—4 дня, если ИЛ-1 удалялся из среды. Увеличение срока инкубации с ИЛ-1 до 2 суток и более приводило к деструкции β-клеток. Ингибиторы NO-синтазы L-NMMA и L-NNA предотвращали повреждение β-клеток, вызванное не только макрофагами, но и цитокинами [938].Эти результаты удалось легко объяснить после того, как выяснилось, что при развитии воспаления экспрессия NO-синтаз наблюдается не только в макрофагах, но также в эндотелиоцитах и β-клетках [490, 903]. В эндотелиальных клетках, полученных из островковых капилляров, выявлена экспрессия как конститутивной, так и индуцибель- ной NO-синтаз, — таким образом, они являются эффективным источником NO-радикалов [1586]. На культурах клеток поджелудочной железы было показано, что ИЛ-ф индуцирует NO-синтазу в β-клетках, но не в α-клетках [490], при этом индукция осуществляется через активацию ядерного фактора транскрипции NF-кВ [960] (рис. 16).
С использованием специфической антисыворотки против макрофагальной NO-синтазы обнаружено, что под действием цитокинов в β-клетках синтезируется аналогичная макрофагальной индуцибельная изоформа фермента [573]. Вместе с тем в β-клетках, как и в эндотелиоцитах, выявляется и конститутивная изоформа NO-синтазы [488]. Главным источником ИЛ-ф при воспалении в поджелудочной железе являются макрофаги и Т- лимфоциты. Совместное действие других провоспалительных цитокинов (ФНО-α и ин- терферон-γ [563] или ФНО-α и липополисахарид [490]) также вызывало индукцию NO- синтазы в островках Лангерганса.
Однако эти цитокины были менее активны, чем ИЛ-ф; более того, индукции NO-синтазы не наблюдалось в выделенных β-клетках (табл. 11). Предполагается, что экспрессия NO-синтазы под влиянием липополисахарида и ФНО-α в β-клетках цельных островков (выявляется иммуногистохимическим методом) происходит посредством активации синтеза ИЛ-ф макрофагами [490].Добавление in vitro к культуре β-клеток соединений, продуцирующих NO·, вызывало гибель клеток, так как они оказались высокочувствительными к действию радикалов [938]. Проведенное сравнительное исследование четырех типов клеток (гепатоциты, активированные и резидентные макрофаги, β-клетки) показало, что β-клетки наиболее подвержены повреждению NO-радикалами, возможно, за счет низкого содержания в них антиоксидантов. В дальнейших исследованиях обнаружено, что гибель β-клеток происходит часто посредством апоптоза.
В низких концентрациях NO-радикалы вызывали изменение синтеза инсулина β-клетками [1572]. Многочисленными исследованиями показано, что индукция цитокинами in vitro NO-синтазы снижает продукцию инсулина в культурах β-клеток, полученных от мышей, крыс и человека [490, 563, 1543, 1744]. Высокая корреляция между образованием NO· и ингибированием секреции инсулина указывает на ключевую роль радикалов в данном процессе, при этом ингибиторы NO-синтазы повышали про-
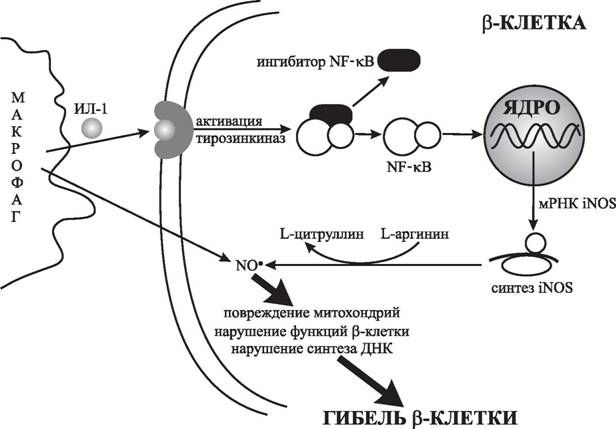
Рис. 16. Механизмы повреждения инсулинсекретирующих клеток макрофагами
Таблица 11
Экспрессия цитокинами и липополисахаридом NO-синтазы в островках Лангерганса и выделенных β-клетках [1127]
| Индуцирующее воздействие (18 часов) | Индукция NO-синтазы (в % от индукции ΗΠ-1β) | |
| Цельные островки | Выделенные β-клетки | |
| Контроль | 0 | 0 |
| ΗΠ-1β (5 ЕД/мл) | 100 | 100 |
| ФНО-α (0,7 нМ) + интерферон-γ (150 ЕД/мл) | 47 ± 18 | Не определялась |
| ФНО-α (0,7 нМ) + липополисахарид (10 мкг/мл) | 50 ± 9 | 0 |
дукцию инсулина, в то время как L-аргинин, напротив, снижал [1474].
Это позволяет предположить, что в организме NO· является одним из регуляторов секреции инсулина поджелудочной железой [1572]. Среди возможных путей снижения синтеза инсулина островковыми клетками может быть ингибирование NO-радикалами аконитазы, участвующей в цикле Кребса [1744]. В то же время инсулин снижает активность индуцибель- ной NO-синтазы и продукцию NO·, возможно, через усиление образования тромбоци- тарного фактора роста [1574].В нарушении секреции инсулина и повреждении островковых клеток участвуют не только NO-радикалы, но и различные токсины, а также другие формы АКМ. Так, диабетом страдает от 30 до 80 % больных гемохроматозом — наследственной патологией, при которой повышено всасывание железа в кишечнике и его накопление в тканях. Будучи эффективным прооксидантом, индуцирующим разложение органических перекисей с образованием реакционных гидроксильных и алкоксильных радикалов, ионы железа могут запускать окислительное повреждение различных клеток и тканей, и в том числе β-клеток поджелудочной железы. На самом деле нельзя утверждать, что какой-то конкретный радикал вызывает деструкцию клеток, так как индукция NO-синтазы вызывается транскрипционным фактором NF-кВ [960], который активируется практически всеми формами АКМ и является медиатором развития окислительного стресса [567]. Прямое измерение методом ЭПР образования липидных радикалов в области поджелудочной железы крыс после введения через желчный проток провоспалительных цитокинов (ФНО-α, ИЛ-ф, интерферон-γ) показало, что главными продуцентами радикалов являются макрофаги и β-клетки, ингибирование индуцибельной циклооксигеназы и транскрипционного фактора NF-кВ снижало образование радикалов [1593]. На свободнорадикальный механизм повреждения β-клеток указывает и тот факт, что токсический эффект NO-радикалов отчасти нивелируется антиоксидантами, в частности α- токоферолом [394, 881], в то же время проведенные на инсулинпродуцирующих клетках RINm5F исследования показали, что в защите от повреждений, индуцированных ИЛ-1 или донорами NO·, γ-токоферол был более эффективен, чем α-токоферол, по-видимому, за счет его высокой способности ингибировать оксиды азота [486, 1530].
При этом островковые клетки поджелудочной железы чрезвычайно уязвимы к свободнорадикальному повреждению, поскольку содержат относительно мало антиоксидантных ферментов. Так, содержание C^Zn-СОД, глутатионпероксидазы и каталазы в β-клетках крысы составляет соответственно 31, 21 и 1 % от концентрации этих ферментов в печени; антиоксидантный дефицит отчасти компенсируется повышенным уровнем ГПО гидроперекисей фосфолипидов [1626]. Аналогичным образом дело обстоит и у мышей, в островках поджелудочной железы экспрессия мРНК C^Zn-СОД составляет 38 % от соответствующего показателя в печени, Mn-СОД — 30 %, глутатионпероксида- зы — 15 %, мРНК каталазы выявлялась в следовых количествах [1006]. Трансфекция генов каталазы и ГПО, позволившая увеличить экспрессию соответствующих ферментов в клетках RINm5F в 100 раз, приводит и к повышению их резистентности к токсическому действию перекиси водорода (в клетках с гиперэкспрессией каталазы — в 10 раз [1626]); индуцированная менадионом (источник О2) или H2O2 гибель клеток мышиной инсулиномы NIT-1, трансфецированных генами Си,/п-СОД и глутатионпероксидазы человека, снижалась в 1,7-2,4 раза [1009].
Еще по теме Роль макрофагов и цитокинов в развитии сахарного диабета типа I:
- Окислительный стресси дисфункция β-клеток при сахарном диабете типа II
- Антиоксиданты в первичной профилактике сахарного диабета
- Антиоксиданты в дебюте сахарного диабета: стратегия вторичной профилактики
- Антиоксиданты в лечении и профилактике сахарного диабета
- Механизмы развития окислительного стресса при диабете типа II
- Сахарный диабет
- Активация макрофагов
- Эмансипация, роль-защитазеленого типа поведения (минус «зеленый»)
- Идеализирование, роль-идолжелтого типа поведения- (плюс «желтый»)
- РОЛЬ-ИДОЛ ТИПА ПОВЕДЕНИЯ (+ "зеленый"): СТРЕМЛЕНИЕ К ЗНАЧИМОСТИ, ТЩЕСЛАВИЕ
- Гармонирование, роль-идол синего типа поведения (плюс «синий»)
- Отчуждение, роль-защита синего типа поведения (минус «синий»)
- РОЛЬ-ЗАЩИТА КРАСНОГО ТИПА ПОВЕДЕНИЯ(минус «красный»): СТРАХ ПЕРЕД ИЗБЫТКОМРАЗДРАЖЕНИЯ И ПЕРЕВОЗБУЖДЕНИЕМ
- 50.Критерии выбора типа стратегического развития организации.
- САХАРНЫЙ ДИАБЕТ
- Нарушение синтеза коллагена. Цитокины и хемокины
- Морализирование, роль-защита красного типа поведения (минус «красный»)
- Характерные изменения при активации макрофагов