Характерные изменения при активации макрофагов
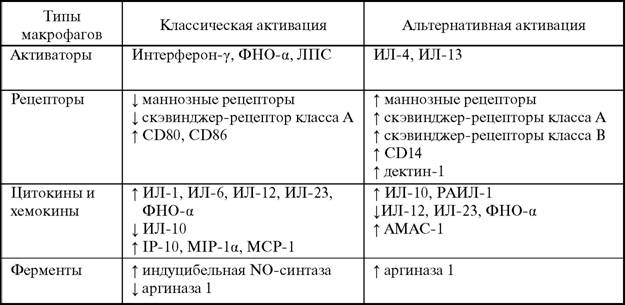
factor-κΒ), вызванную ФНО-α [1036]. Инкубация в течение 16 часов человеческих моноцитов с ИЛ-4 и М-КСФ в 6—12 раз повышала, а с ЛПС и дексаметазоном — снижала содержание в клетках мРНК CD36, в то время как интерферон-γ, ФНО-α и ИЛ-3 не влияли на экспрессию CD36 [1813].
В культурах клеток RAW 264.7 экспрессия CD36 увеличивалась в ответ действие окисленных липопротеинов и ИЛ-4 и зависела от протеинкиназы С (PKC) и ядерного гормонального фактора транскрипции PPAR-γ [599]. Важным рецептором для связывания β-глюканов (структурный компонент грибковых клеток) является дектин-1. В мышиных макрофагах его экспрессия возрастала в ответ на действие ИЛ-4, ИЛ-13, TGF-β и ГМ-КСФ, но снижалась при инкубации клеток с ЛПС, ИЛ-10 и интерфероном-γ [1765].В ходе активации макрофагов по классическому пути происходит усиление продукции провоспалительных цитокинов, таких как ФНО-α, ИЛ-1, ИЛ-6 и ИЛ-12, которые могут включаться в аутокринную регуляцию [1175]. Наличие аутокринной регуляции позволяет усилить ответ клеток на воздействие и сделать его ответ менее специфичным. В результате этого клетки на разные действующие стимулы отвечают характерным однонаправленным изменением функциональных показателей, что необходимо для развития типового патологического процесса. Напротив, при альтернативной активации повышается синтез антивоспалительных цитокинов, таких как ИЛ-10, рецепторных антагонистов ИЛ-1 (РАИЛ-1) и хемокинов (АМАС-1) [678]. Связываясь со специфическим рецептором для ИЛ-ф, РАИЛ-1 блокирует действие цитокина и защищает клетки от токсического действия активных форм кислорода [62]. Соотношение цитокинов ИЛ- ф/РАИЛ-1 имеет важное значение для развития многих воспалительных процессов [56, 271]. Повышенная экспрессия РАИЛ-1 у содержащихся на гиперхолестериновой диете апо Е-дефицитных мышей в среднем на 50 % снижала формирование атеросклеротических бляшек, в аналогичных условиях у нокаутов по РАИЛ-1 наблюдались массивные воспалительные инфильтраты, которые часто приводили к гибели животных [1139].
В общем случае можно сказать, что классическая активация макрофагов на начальных стадиях воспаления сопровождается повышением их микробицидной и туморицидной активности, в то время как альтернативная активация на более поздних стадиях ведет к снижению цитотоксического действия и усилению эндоцитоза и фагоцитоза продуктов клеточной деструкции [1339].Многие исследователи наиболее важным критерием различия данных путей активации считают соотношение интенсивности катаболизма L-аргинина через NO- синтазу и аргиназу (рис. 3). В этой связи определение ферментативной активности

Рис. 3. Метаболизм L-аргинина под действием цитокинов, бактерий и глюкокортикоидов. ОДК — орнитиндекарбоксилаза; ОАТ — орнитинаминотрансфераза
NO-синтазы и аргиназы часто используется в качестве показателя поляризации макрофагов и их активации по классическому или альтернативному пути [831, 1179]. Усиление транскрипции мРНК индуцибельной NO-синтазы происходит в ответ на действие многих провоспалительных цитокинов, в то время как стероиды, TGF-β, р53 и сам NO· снижают активность фермента [1379]. Внутриперитонеальное введение мышам казеина, каррагинана, вакцин БЦЖ или вируса болезни Ньюкастла через 24 часа повышало содержание индуцибельной NO-синтазы в перитонеальных макрофагах в 10-25 раз [778].
Сегодня практически во всех работах по активации макрофагов высказывается не совсем верное утверждение о том, что при классической активации возрастает продукция цитотоксических радикалов NO· и O2 [1093, 1175]. Если для анализа продукции NO· и O- применяются неспецифические индикаторы, то в таких исследованиях при активации макрофагов обнаруживается увеличение продукции обоих радикалов, однако специфические метки на NO· и O2- выявляют реципрокные взаимоотношения между этими молекулами [129]. Дело в том, что в макрофагах и во многих других клетках радикалы NO· и O- образуют регуляторный элемент, в котором увеличение продукции одной молекулы автоматически сопровождается снижением концентрации другой [147].
В нормальных условиях в клетках работают кальций- и кальмодулинзависимые конститутивно экспрессируемые NO-синтазы, которые создают достаточно постоянные невысокие концентрации NO·. Основным назначением данных ферментов является регуляция тонуса сосудов, нейротрансмиссия, а также контроль работы дыхательных цепей митохондрий. Действие на клетки микроорганизмов, бактериальных липополисахаридов и целого ряда других факторов приводит к активации мембранных NADPH-оксидаз и ксантиноксидазы, что индуцирует быструю и интенсивную продукцию супероксид- аниона. Радикалы NO· и O2- эффективно взаимодействуют между собой с образованием пероксинитрита, как уже отмечалось, константа скорости этой реакции определяется скоростью диффузии молекул и приближается к 1010 М-1с-1 [147]. В результате такого взаимодействия базальная концентрация NO· снижается, что ведет к вазоконстрикции, агрегации тромбоцитов и адгезии нейтрофилов. Увеличение экспрессии индуцибельной NO-синтазы сопровождается значительным повышением продукции NO· (приблизительно в 100 раз) и снижением внутриклеточной концентрации супероксидного анион-радикала.В клетках млекопитающих выявляется два типа аргиназ: цитозольная аргиназа 1 и митохондриальная аргиназа 2. Хотя изоформы фермента катализируют одну реакцию, они кодируются разными генами, отличаются по структуре и клеточной локализации. В печени аргиназа 1 служит для катаболизма аминогрупп посредством перевода их в мочевину и выведения из организма. В клетках других тканей (макрофагах, гладкомышечных клетках, эндотелиоцитах) роль аргиназы остается неясной [289]. Считается, что макрофагальная аргиназа 1 отвечает за синтез полиаминов и пролина, которые необходимы для клеточной пролиферации, в том числе фибробластов, и продукции коллагена. Под действием цитокинов второго типа ИЛ-4, ИЛ-13 и TGF-β в макрофагах повышается экспрессия аргиназы 1, но не аргиназы 2 [298, 674]. Напротив, ИЛ-10 увеличивал активность аргиназы 2, но не влиял на активность аргиназы 1 [298].
Инкубация перитонеальных макрофагов мыши с ИЛ-4 (10 нг/мл) повышала транскрипцию гена аргиназы 1 в 88 раз, что естественно, приводило к снижению концентрации аргинина и продукции NO· [1742]. Как правило, индуцирующие действия цитокинов первого и второго типа на NO-синтазу и аргиназу находятся в реципрокных взаимоотношениях, т. е. активация одного фермента сопровождается ингибированием другого [1159]. В частности, L-гидроксиаргинин, промежуточных продукт катализируемой NO-синтазой реакции, прямо ингибирует в клетках аргиназу 1 [750], а под действием ИЛ-4 и ИЛ-13 в мышиных макрофагах независимо от аргиназы снижается активность индуцибельной NO-синтазы [674]. При этом как классический, так и альтернативный пути активации макрофагов ведут к усилению транспорта L-аргинина в клетки через катионный аминокислотный транспортер САТ2, что повышает эффективность его метаболизма либо по NO-синтазному, либо по аргиназному механизму [1812].Функциональное состояние клеток во многом зависит от окислительно-восстановительного баланса, который в свою очередь определяется соотношением восстановленного и окисленного глутатиона. Показано, что состояние М1 характеризуется высоким содержанием глутатиона и высоким уровнем его восстановленности [1181]. Так, при активации мышиных макрофагов (линия RAW 264.7) липополисахаридом и/или интерфе- роном-γ концентрация внутриклеточного глутатиона возрастала в 2—5 раз [602]. Предполагается, что глутатион необходим для перевода NO-синтазы в активное состояние и его содержание тесно коррелирует с синтезом NO· макрофагами [388]. Кроме того, высокий уровень глутатиона снижает индуцированный донорами NO· и H2O2 апоптоз макрофагов [348]. При альтернативной активации состояние М2 характеризуется сниженным уровнем глутатиона и увеличенным соотношением GSSG/GSH [1181].
В организме человека функциональное состояние макрофагов должно находиться под строгим контролем. Нарушение такого контроля может привести к развитию опасного синдрома активации макрофагов, при котором наблюдаются повреждения многих органов [1383].
Синдром активации макрофагов — редкое, но тяжелое состояние, зачастую фатальное (при его развитии погибает от 20 до 60 % пациентов). Возникает оно, как правило, вторично в результате иммунной гиперактивации, сопровождающей системные инфекции, аутоиммунные расстройства, злокачественный рост, и характеризуется лимфоаденопатией, панцитопенией, гепатоспленомегалией, печеночной недостаточностью, коагулопатией, неремиттирующей лихорадкой, гипертриглицери- демией, гиперферритинемией; существуют также первичные, наследственные формы заболевания [334, 954]. Синдром активации макрофагов связан с высокой пролиферативной активностью Т-лимфоцитов и метаболической активности макрофагов, в крови больных наблюдается высокое содержание провоспалительных цитокинов ИЛ-ф, ФНО-α, ИЛ-6, ИЛ-18 и интерферона-γ. В клинических условиях при развитии синдрома активации макрофагов необходима быстрая терапия глюкокортикоидами и имму- носупрессирующими препаратами, снижающими выработку провоспалительных цитокинов [954, 1383].