ОКИСЛИТЕЛЬНЫЙ СТРЕСС И СТАРЕНИЕ
Взять человека рыжего и веснушчатого, кормить плодами до 30 лет, а затем опустить в каменный сосуд с медом и закупорить. Через 120 лет употреблять вовнутрь. Древнеперсидский рецепт вечной молодости
Биология преподносит немало удивительных примеров того, что некоторые живые существа, казалось бы, вообще не подвержены старению и «естественной смерти».
Близ Балеарских островов на дне Средиземного моря найдено водное растение посидония (Posidonia oceanica), чьи стебли простираются примерно на 8 километров, по оценкам специалистов возраст данного экземпляра посидонии достигает примерно 100 тыс. лет. Найденный в лесах Тасмании 10 лет назад клон кустарника остролиста королевского (Lomatia tasmanica), размножающегося корневыми побегами, живет уже более 43 тыс. лет. Есть деревья, возраст которых составляет несколько тысяч лет, и они продолжают плодоносить. Так, в Северной Америке обнаружены деревья сосны остистой (Pinus longaeva) и секвойи гигантской (Sequoia gigantea) в возрасте более 4000 и 2500 лет соответственно. В Южной Америке к долгожителям (около 2000 лет) относится фицройя (Fizroya cupressoides), а в горах Тянь-Шаня находят 1300-летние деревья арчи туркестанской (Juniperus turkestanica). При этом высота калифорнийской секвойи, занесенной в Книгу рекордов Гиннеса за свой рост, может достигать 113 метров. Некоторые виды морских черепах, двустворчатых моллюсков и омаров, возможно, не подвержены старению или утратили эту способность; в октябре 2007 г. у берегов Исландии обнаружен моллюск Arctica islandica, получивший имя Мин в честь китайской императорской династии, в период правления которой он родился — 405—410 лет назад, это абсолютный верифицированный рекорд для животных. У черепах возраст, превышающий 150 лет, не является предельным, причем животные и в этих преклонных годах часто не обнаруживают биологических признаков старения. То же относится к некоторым видам рыб и птиц, для которых не наблюдается возрастного снижения темпа воспроизводства, физических способностей и устойчивости к заболеваниям.Среди многоклеточных животных потенциально бессмертной является гидра (Cnidaria vulgaris), которая относится к кишечнополостным [28]. Гидра имеет простое строение и представляет собой трубку, сформированную из примерно 20 клеточных типов, являющихся продуктами дифференцировки трех линий (двух эпителиальных и интерстициальной) стволовых клеток. Эпителиальные клетки стенки тела непрерывно пребывают в митотическом цикле и как бы текут с середины тела к концам, постоянно перемещаясь или вверх к голове, или к подошве. Неделящиеся дифференцированные клетки замещаются в теле гидры в течение 20 дней. Связанные с возрастом изменения в клетках предотвращаются их быстрой сменой на новые. При неблагоприятном изменении температуры или состава воды деление замедляется и гидра стареет, т. е. в зависимости от внешней среды потенциально нестареющий организм может переходить в стареющее состояние.
В сообществе млекопитающих бессмертных нет. Если они не погибают от внешних причин до наступления старости, то умирают от болезней, связанных с возрастной дряхлостью. Как ни странно, человек относится к долгожителям, и по продолжительности жизни, а также по способности сохранять высокую физическую и интеллектуальную активность в преклонном возрасте может сравниться с самыми долгоживущими рыбами, рептилиями и птицами. Действительно, средняя продолжительность жизни приблизилась в развитых странах к 80 годам. Если доверять только строго документированным данным, максимальная продолжительность жизни (максимальный возраст, до которого наблюдалось доживание представителей данного вида) у человека составляет 120 лет. Рекордсменом (занесена в Книгу рекордов Гиннеса) считается француженка
Жанна Кальмон, которая прожила 122 года и 164 дня. Старость обычно ассоциируется с неизбежной деградацией физического и психического здоровья человека. Но в ряде исследований показано, что среди «глубоких стариков» совсем немало тех, кто сохраняет хорошее здоровье, высокую работоспособность, удивительную творческую активность.
Научные исследования явлений старения и долгожительства связаны с надеждой, что их результаты помогут человеку избавиться от дряхлости, а может быть, и откроют пути повышения верхней границы продолжительности жизни человека.В XIX столетии французский естествоиспытатель Жан-Батист Ламарк впервые ввел понятия случайной смерти в результате болезней, убийства или каких-либо несчастных случаев и естественной смерти, вызванной старением организма. Он постулировал, что естественная смерть вызывается внутренними причинами, лежащими в основе старения. Наука, изучающая феномен старения живых существ, в том числе человека, получила название «геронтология» (от греч. γεροντ (род. падеж γεροντος) — «старец» и λόγος — «мысль», «учение»). Этот термин был впервые предложен И. И. Мечниковым в 1903 г. Современная геронтология — междисциплинарная наука, в состав которой входят биология старения, клиническая геронтология (гериатрия), геронтопсихология и социальная геронтология (геронтогигиена). Фундаментальное значение геронтологии определяется направленностью на изучение наиболее существенных вопросов жизни, а именно ее продолжительности и смерти. Эти вопросы важны как для каждого отдельного индивидуума, так и всего человечества. Согласно современным представлениям под старением понимают приходящее с возрастом замедление, угнетение и потерю физиологических функций организма, сопровождающееся повышенной частотой онкологических и дегенеративных заболеваний.
На сегодняшний день предложено более 300 теорий, объясняющих феномен старения, что делает проблемой даже их классификацию [1738]. Почти все теории сводятся к вариациям двух концепций: 1) старение — это генетически запрограммированный процесс, необходимый для быстрой смены поколений в популяции и эволюционного развития; 2) старение — это стохастический, случайный процесс, обусловленный «изнашиванием» организма в результате самоотравления продуктами жизнедеятельности и/или повреждения, наносимого постоянно действующими вредными факторами внешней среды.
Наиболее весомым аргументом в пользу первой концепции является ограниченное количество делений, которые способны совершать соматические клетки («лимит Хейфлика»). В качестве механизма отслеживания количества делений предлагается уменьшение длины концевых участков хромосом — теломер. Концепция самоотравления организма впервые предложена Мечниковым, который считал толстый кишечник своего рода инкубатором для гнилостных микроорганизмов, продукты обмена которых отравляют организм, тем самым сокращая продолжительность жизни. Сегодня многочисленные теории «накопления повреждений» связывают с истощением механизмов репарации повреждений, вызванных свободными радикалами, окислением, гликозилированием и т. д. Обе концепции приводят к единому итогу: в результате старения повреждается определенный клон клеток, как следствие возникают нарушения нейроэндокринной и иммунной регуляции, снижаются адаптационные возможности организма, что лежит в основе развития старческих патологий, приводящих к летальному исходу.Впервые свободнорадикальная теория старения организмов была предложена в 1956 г. Денхамом Харманом, который обратил внимание на сходство патологических изменений, таких как мутагенез, канцерогенез, массивная клеточная гибель, наблюдаемых в процессе старения организма и при действии ионизирующей радиации [716]. К тому времени уже было известно, что при действии радиации образуются ОН-ради- калы, а методом ЭПР было показано наличие радикалов в животных тканях. Харман предположил, что образующиеся в живых клетках и тканях радикалы вызывают повреждения, приводящие к старению организма. На сегодняшний день выполнено значительное число исследований и получено много данных в пользу такой теории старения, однако каких-либо однозначно интерпретируемых результатов получено не было. Общий анализ данных об участии АКМ в процессах старения и сопровождающих старение заболеваний позволяет ряду авторов утверждать, что повреждение под действием АКМ макромолекул приводит к мутациям, нестабильности генома в целом и развитию ряда возрастных патологий: рак, сердечно-сосудистые заболевания, возрастная иммунодепрессия, дисфункция мозга, остеопороз, катаракта и др.
Следует отметить, что все «старческие хвори» (болезни Альцгеймера, Паркинсона, инфаркт миокарда, диабет типа II, остеоартриты и ревматоидные артриты) связаны с развитием окислительного стресса (см. с. 9—10). Свободнорадикальная теория старения достаточно логично объясняет возрастное повышение частоты развития онкологических патологий: повреждающие воздействия АКМ на геном приводят либо к опухолевой трансформации клеток, либо к их апоптозу, что в конечном счете ведет к старению организма [1310].Конкретный механизм индукции старения свободными радикалами сегодня мало понятен. Предполагают, что АКМ повреждают хроматин и ДНК, жизненно важные белки, мембраны, коллаген, изменяют регуляцию внутриклеточного кальция и пр. C возрастом повышается содержание в клетках окислительно поврежденных биомолекул, F2-изопростанов в сыворотке и органах (печень, почки, мышцы) [1722]. Карбонильные группы (С=О) в белках образуются при взаимодействии с гидроксиноне- налями или катализируемом ионами металлов окислении аминокислотных остатков пролина, аргинина, лизина, треонина или гистидина. Карбонилирование белков ускоряет их деградацию протеасомами, что необходимо для удаления функционально нарушенных белков [1225], но вместе с тем приводит к образованию белковых агрегатов, которые снижают активность протеасом. В физиологических условиях может быть кар- бонилировано до 10 % белковых молекул, с возрастом содержание модифицированных протеинов в фибробластах, хрусталике глаз, клетках головного мозга человека повышается [1012, 1174]. При этом наибольшее накопление агрегированных карбонилирован- ных белков выявляется в клетках с низкой пролиферативной активностью, в частности в нервных клетках. С возрастом также повышается содержание карбонильных групп в сывороточных белках альбумине, трансферрине и aj-макроглобулине [816].
Поскольку более 90 % всего кислорода, потребляемого клетками аэробного организма, утилизируется митохондриями, то можно ожидать, что основное количество O2 и H2O2 образуется именно в митохондриях.
В 1972 г. Харман высказал предложение, что именно в этих органеллах находится счетчик биологического времени [720]. Идею быстро подхватили, и митохондрии даже были названы «ахиллесовой пятой клетки». Показано накопление в митохондриях карбонильных групп в составе адениннуклео- тидтранслоказы и аконитазы [1796, 1797], сопровождающееся падением активности этих ферментов. Снижение интенсивности переноса электронов в дыхательной цепи митохондрий с возрастом в первую очередь определяется уменьшением активности комплексов I и IV (соответственно NADH-убихинолредуктазы и цитохромоксидазы). Комплексы II и III более устойчивы как к окислению, так и к старению. С возрастом также отмечается значительное падение активности митохондриальной NO-синтазы (табл. 19). Анализ показывает, что продукция АКМ митохондриями долгоживущих млекопитающих и птиц значительно меньше, чем у представителей короткоживущих видов [378, 1456], при этом снижение калорийности питания увеличивает продолжительность жизни и угнетает продукцию АКМ в митохондриях. Аналогичные результаты получены у животных с гиперэкспрессией разобщающего белка UCP2, имеющих низкую температуру тела и повышенную продолжительность жизни (в среднем на 15 %) [483].В большинстве соматических клеток млекопитающих митохондрии являются главным продуцентом АКМ, поэтому вполне естественно рассматривать их в качестве «пружины» биологических часов. Хотя появление восстановленных форм кислорода (O2 и H2O2) показано на разных участках цепи переноса электронов, большинство исследователей основным источником O2 в митохондриях считают цикл убихинона [147]. Естественно, что содержание убихинона должно сказываться и на внутриклеточной
Таблица 19
Содержание ТБК-РП, карбонильных групп и активность ферментов в митохондриях мозга мышей разного возраста [1201]
| 28 недель | 52 недели | 78 недель | |
| ТБК-РП | 5,2 ± 0,4 | 8,1 ± 0,4 | 9,1 ± 0,4 |
| Карбонильные группы в белках | 55 ± 4 | 74 ± 5 | 91 ± 5 |
| NO-синтаза | 0,59 ± 0,06 | 0,33 ± 0,04 | 0,16 ± 0,02 |
| NADH-цитохром-с-редуктаза | 319 ± 10 | 270 ± 10 | 210 ± 10 |
| Цитохромоксидаза | 116 ± 8 | 90 ± 7 | 75 ± 8 |
| При содержании на обогащенной витамином Е диете (4,3 г токоферилацетата/кг корма) | |||
| ТБК-РП | 5,9 ± 0,3 | 5,5 ± 0,4 | 5,9 ± 0,4 |
| Карбонильные группы в белках | 50 ± 5 | 59 ± 4 | 63 ± 5 |
| NO-синтаза | 0,62 ± 0,05 | 0,54 ± 0,05 | 0,43 ± 0,06 |
| NADH-цитохром-с-редуктаза | 322 ± 11 | 290 ± 10 | 283 ± 11 |
| Цитохромоксидаза | 120 ± 8 | 112 ± 8 | 102 ± 8 |
продукции АКМ, и на продолжительности жизни. Группой американских ученых было проведено исследование длительного (начиная с 3 месяцев и до конца жизни) содержания мышей линии С57BL/6 на диете, обогащенной ^Q10 (ежедневные дозы составляли 93 и 371 мг/кг массы тела) [1540]. У экспериментальных животных повышалось содержание ^Q9 и ^Q10 в гомогенатах и митохондриях печени, сердца, почек и скелетных мышц, но не изменялось в гомогенатах мозга. Не было выявлено однозначного изменения продукции O2 и содержания карбонильных групп в белках митохондрий, продолжительность жизни мышей, получающих ^Q10, также не отличалась от контрольной группы. В другом исследовании, выполненном в Институте биохимической физики РАН им. Н. М. Эмануэля, ^Q10 в дозе 0,4 мг/кг массы тела незначительно снижал продолжительность жизни мышей линии СВА [159]. Такой эффект ^Q10 может быть обусловлен тем, что большая часть митохондриального убихинона не связана с электронтранспортными белками, поэтому эффективно не участвует ни в продукции O2, ни в защите от повреждающего действия АКМ [650]. Положительный эффект увеличения средней продолжительности жизни на 11,7 % и максимальной продолжительности жизни на 24 % для ^Q10 (0,7 мг/кг в день) был получен на самцах крыс линии Вистар, которые содержались на обогащенной ненасыщенными жирными кислотами диете [1365].
Продукция H2O2 митохондриями в значительной степени определяется наличием белка p66Shc (src homology 2 domain-containing transforming protein C1), который способен переносить электроны с электронтранспортной цепи через цитохром с на молекулярный кислород [662]. В физиологических условиях белок p66Shc отвечает за усиление продукции H2O2 в ответ на стрессовые воздействия и факторы роста. Считается, что около 30 % внутриклеточной H2O2 синтезируется через p66Shc. В экспериментах, выполненных на трансгенных мышах, показано, что выключение гена, кодирующего p66Shc, приводит к увеличению продолжительности жизни на 30 % [1146] и снижению проявлений ассоциированных с возрастом патологий: предотвращает АКМ-зависимую дисфункцию эндотелия у старых животных [617], способствует существенному снижению выраженности окислительного стресса и проявлений гломерулопатии при стрептозо- тоцин-индуцированном диабете [1138], а также препятствует атерогенезу (содержание на высокожировой диете гомозиготных мышей p66Shc-/- сопровождалось увеличением площади атеросклеротического поражения их аорты лишь на 3 %, в то время как мышей дикого типа — на 21 %) [1197]. У млекопитающих p66Shc является первым геном, выключение которого повышало продолжительность жизни [1147].
Многие современные теории старения предполагают, что ДНК служит основной мишенью действия эндогенных и экзогенных повреждающих агентов в клетке. Повреждения ядерной и митохондриальной ДНК соматических клеток, такие как точечные мутации, делеции и транслокации, приводят к активации или инактивации специфических генов, вовлеченных в регуляцию клеточного цикла и контроль роста. Накопление с возрастом таких мутаций в различных органах и тканях является основным фактором, определяющим развитие возрастных патологий, включая рак. У экспериментальных животных (мыши и крысы) с возрастом в ДНК из разных органов (печень, сердце, почки, мышцы, головной мозг) содержание 8-оксо-2'-дезоксигуанозиновых оснований увеличивается в 2—5 раз, наибольшее количество окисленных оснований выявляется в митохондриальной ДНК [708, 782, 1412]. Это позволяет предположить, что мутации и снижение транскрипционной активности митохондриальной ДНК являются важным элементом клеточного старения. У мышей, дефицитных по полимеразе-γ, повышен уровень образования мутаций в ДНК митохондрий и значительно снижена продолжительность жизни [936]. Функциональные нарушения митохондрий ассоциируются с основными возрастными патологиями: атеросклерозом [1069], нейродегенеративными заболеваниями [1705], диабетом [1424], опухолевыми процессами [1712].
Особенности окислительного повреждения ДНК позволяют найти точки соприкосновения свободнорадикальной и теломерной теориям старения. Согласно теломерной теории, вследствие особенностей репликативного аппарата зрелые соматические клетки взрослых индивидуумов могут делиться ограниченное число раз («лимит Хейф- лика»), поскольку каждый акт удвоения хромосом приводит к укорочению ДНК на 20—200 пар оснований, при этом недореплицируются теломеры — особые некодирующие концевые участки ДНК длиной 4—15 тысяч пар оснований (у человека), представленные повторами гексануклеотидов 5'-TTAGGG-3' [1142]. Таким образом, через определенное число делений клетки ее теломеры укорачиваются до некоего предела, после чего происходит апоптоз «постаревшей» клетки или необратимая остановка роста. Транспонируя теломерную теорию старения на целый организм, можно привести свидетельствующие в пользу ее справедливости факты: так, теломеры в клетках печени стариков старше 80 лет почти вдвое короче, чем у детей до 8 лет [1599].
Укорачиванию теломер в процессе жизнедеятельности организма способствует не только концевая недорепликация, но и их окислительное повреждение [775]. Повышенной чувствительностью к окислительному стрессу концевые участки хромосом обладают в силу высокого содержания гуаниновых оснований. Более того, в составе теломер остатки гуанина окисляются более интенсивно, чем в других участках ДНК: так, после облучении УФ-светом образование 8-оксо-7,8-дигидро-2'-дезоксигуанозина в теломерных последовательностях [5'-CGC(TTAGGG)7CGC-3'] было в 5 раз выше, чем в нетеломерных [5'-CGC(TGTGAG)7CGC-3'], — очевидно, в силу высокой уязвимости центрального остатка гуанина в триплетах GGG, поскольку образующийся в ходе его окисления промежуточный продукт катион-радикал гуанина наиболее стабилен [1237]. еще одной причиной ускорения укорачивания теломер под действием АКМ служит меньшая эффективность репарации концевых участков хромосом по сравнению с другими участками ДНК. Так, показано, что после 45-минутной обработки Н2О2 (300 мкМ) культур фибробластов MRC-5, WI-38, F15 и F16 в их теломерных фрагментах однонитевые разрывы не только возникали в большем количестве (в среднем в 3 раза), чем в мини-сателлитах или кодирующих последовательностях, но и они хуже репарировались: даже через 19 дней в теломерах оставалось от 18 до 76 % (в зависимости от культуры клеток) повреждений, в то время как в других фрагментах ДНК уже через 24 ч однонитевые разрывы были полностью удалены [1314].
С возрастом повышается содержание окисленных белков: у старых животных около половины всех внутриклеточных белков окисленно модифицированы. Хотя окислению могут подвергаться все белки, многие из них имеют критические чувствительные к действию АКМ элементы, окислительная модификация которых может обратимо или необратимо изменять функциональное состояние белка. Чувствительны к окислению и нитрозилированию кальцийрегулирующий белок кальмодулин и кальциевые каналы, их альтерация приводит к нарушению кальциевого гомеостаза [1548]. Интересно отметить, что в процессе постнатального развития человека в структурах головного мозга выявлены разнонаправленные изменения содержания продуктов ПОЛ и окислительной модификации белков [42]. Во всех отделах головного мозга зарегистрировано возрастное увеличение уровня продуктов окислительной модификации белков, достигающее максимума к 12—21 годам, при этом наиболее выраженное (4—6-кратное) повышение наблюдалось в зрительной коре, гиппокампе, диэнцефальных и понтобульбарных отделах головного мозга. Отчетливое онтогенетическое снижение содержания продуктов ПОЛ отмечено в коре поля 17, структурах «древней коры» и в гипоталамусе.
В составе белков наиболее подвержены окислению серосодержащие аминокислотные остатки цистеин и метионин. Обратимое окисление/восстановление метионина проходит через образование метионинсульфоксида, для восстановления которого служит метионинсульфоксидредуктаза (Msr) (КФ 1.8.4.6, систематическое название — протеин^-метионин:тиоредоксин-дисульфид^-оксидоредуктаза), а донором атомов водорода выступает тиоредоксин (рис. 29). Поскольку при окислении образуются S- и R-изомеры метионинсульфоксида, многие организмы содержат два класса метионин- сульфоксидредуктаз (MsrA и MsrB), восстанавливающих соответственно S- и R-изомеры. C возрастом с 10 до 80 лет в белках хрусталика глаз и коллагене кожи человека соотношение метионинсульфоксид/метионин увеличивается в 3—4 раза [1551]. В белках из мембран эритроцитов содержание метионинсульфоксида с возрастом повышается на
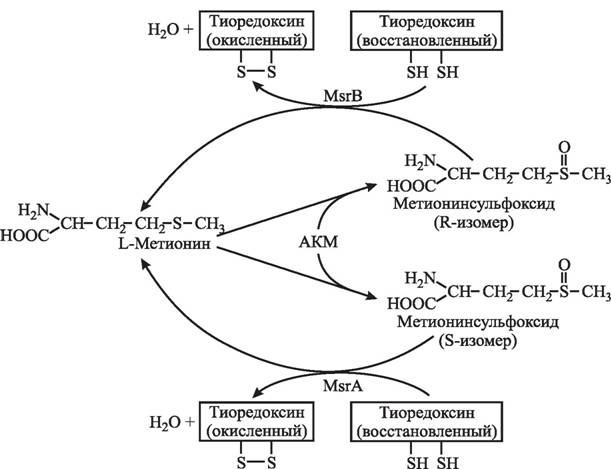
Рис. 29. Циклические окислительно-восстановительные преобразования метионина [1551]
50—60 % [1488]. В тканях мутантных по MsrA мышей наблюдалось повышенное содержание карбонильных групп, эти животные были менее устойчивыми к действию гипер- оксии (100 % О2), продолжительность жизни у них была снижена на 40 % по сравнению с мышами исходной линии [1173]. Гиперэкспрессия MsrA у Drosophila melanogaster приводила к увеличению продолжительности жизни в среднем на 70 % [1433]. Прямая зависимость продолжительности жизни от активности MsrA и MsrB характерна для клеток дрожжей [914]. Восстановителем для метионинсульфоксидредуктаз выступает тиоредоксин, который, в свою очередь, восстанавливается тиоредоксинредуктазой с участием NADPH [147].
Другой предпочтительной для окисления мишенью в белках являются ОН-группы серина, треонина и тирозина. Их фосфорилирование и дефосфорилирование в составе белков соответственно протеинкиназами и протеинфосфатазами является важной сигнальной системой, служащей для регуляции активности ферментов в клетках и передачи внешних сигналов на геном. Особенно это касается тирозиновых остатков, окисление или нитрозилирование которых нарушает их способность фосфорилироваться тирозинкиназами и, соответственно, участие в работе данной сигнальной системы, отвечающей на действие ряда факторов роста и гормонов [323]. С тирозиновыми остатками эффективно взаимодействует пероксинитрит, который в присутствии СО2 переходит в метаболиты O2=NOOCO2 или O2NOCO2, нитрозилирующие ароматические аминокислоты [323]. C возрастом увеличивается содержание 3-нитротирозина в белках [556], что сопровождается снижением активности ряда ферментов. В частности, в клетках скелетных мышц старых крыс значительно снижена активность гликогенфосфорилазы b в результате нитрования входящих в ее состав тирозиновых остатков [1497]
Характерным показателем старения организма служит накопление в клетках и тканях жиробелковых пигментов липофусцина и цероида. Эти пигменты старения имеют практически одинаковые физико-химические и гистохимические свойства. В морфологии их различают по локализации. Липофусцин (от греч. λίπος — «жир» и лат. fus- cus — «темный, бурый») — мелкий гранулярный золотисто-коричневый пигмент, образованный из фосфолипидов и белков. Образуется липофусцин в результате сшивок белковых и липидных молекул малоновым диальдегидом, он накапливается в виде мелких желто-бурых глыбок в цитоплазме клеток всех органов и тканей, нарушая их функции. Содержание липофусцина увеличивается при старении организма и тяжелых истощающих заболеваниях с атрофией органов. В клетках мозга, сердца, кожи и др. накопление липофусцина в старческом возрасте может достигать 30 % их объема. При некоторых формах энцефалопатий липофусцин может занимать до 80 % объема нервных клеток. Цероид — липопигмент, образующийся в макрофагах путем гетерофагии при резорбции липидов. К образованию гетерофагических вакуолей (липофагосом) приводит эндоцитоз. Липофагосомы трансформируются во вторичные лизосомы (липофаголизосомы). В них липиды частично перевариваются лизосомными ферментами и остаются внутри. Это так называемые третичные фаголизомы, или телолизосомы, которые и содержат вещество под названием цероид. В условиях патологии образование цероида чаще всего отмечается при некрозе тканей, особенно в участках кровоизлияний.
Выше мы рассматривали главным образом накопление повреждений, связанных с продукцией АКМ. С возрастом функциональные изменения могут быть связаны не только с повреждением биомолекул, но и с нарушением регуляторной роли АКМ. Важной функцией NO-радикалов является регуляция тонуса сосудов в ответ на действие вазодилататоров, сдвиговое напряжение и другие факторы [147]. C возрастом снижается расширение сосудов в ответ на действие ацетилхолина. Так, если максимальную аце- тилхолин-индуцированную релаксацию аорты молодых (4—6 недель со дня рождения) крыс принять за 100 %, то у взрослых животных (3—6 месяцев) она составляет 50 % и лишь 25 % — у старых (12—25 месяцев) крыс [1116]. Возрастные нарушения релаксиру- ющей функции эндотелия артерий могут быть связаны с активацией ксантиноксидазы, продуцирующей О2, который, в свою очередь, ингибирует NO· [1205]. Течение типовых патологических процессов, прежде всего воспаления, связано с активацией редокс- чувствительных факторов транскрипции, поэтому изменения регуляторной роли АКМ могут явиться причиной развития хронических патологий. Так как многие возрастные патологии, такие как слабоумие, рак, сердечно-сосудистые заболевания, артриты, связаны с активацией провоспалительных генов, то можно предположить, что в основе их развития могут лежать нарушения редокс-регуляции. И действительно, все эти заболевания в той или иной степени связаны с высокой активностью микроглии и макрофагов. Следствием таких рассуждений является «воспалительная теория» старения [465].
Хотя повреждения биомолекул наблюдаются во всех органах, однако устойчивость отдельных клеток и тканей к таким повреждениям разная. Можно выделить наиболее чувствительные к окислительным повреждениям группы клеток, функциональные нарушения в которых лежат в основе развития широкой группы старческих патологий. В группу старческих патологий входят нейродегенеративные патологии, диабет, онкологические патологии. По данным американских исследований, в 2000 г. в США насчитывалось 14,2 млн больных диабетом, 4,5 млн людей с болезнью Альцгеймера и около 1 млн — с болезнью Паркинсона [1712].
Еще по теме ОКИСЛИТЕЛЬНЫЙ СТРЕСС И СТАРЕНИЕ:
- Механизмы развития окислительного стресса при диабете типа II
- Окислительный стресс и инсулинрезистентность
- Окислительный стресс и диабетические ангиопатии
- Окислительный стресси дисфункция β-клеток при сахарном диабете типа II
- Меньщикова Е. Б.. Окислительный стресс: Патологические состояния и заболевания / Е. Б. Меньщикова, Н. К. Зенков, В. З. Ланкин, И. А. Бондарь, В. А. Труфакин.— Новосибирск,2008. - 284 с., 2008
- Психология травматического стресса. Методологические различия при определении стресса и травматического стресса
- Биологические подходы к старению
- Показатели биологического возраста (маркеры старения)
- Окислительная модификация ЛНП и атеросклероз
- Экспериментальные модели окислительного поражения легких у животных
- Личность и старение
- Глава первая Загадки старения — слово науке
- Системное изменение активности свободнорадикальных окислительных процессов
- Б. Электронные влияния на окислительно-восстановительные (редокс) потенциалы.
- О.В.Краснова. Психология старости и старения, 2003
- Физические аспекты старения и проблема здоровья
- Старение населения и его влияние на оказание помощи в финальном периоде жизни - важнейшие вопросы общественного здравоохранения в XXI веке