Ферментативные антиоксиданты
На сегодняшний день накопилось много свидетельств в пользу свободнорадикальной теории старения. Исследование антиоксидантного статуса у людей в разных возрастных группах (35—39 лет, 50—54 года, 65—69 лет) не выявило существенных различий в содержании СОД, глутатиона, каталазы и церулоплазмина [300].
Наиболее весомым аргументом является сильная корреляция между выраженностью окислительного стресса и продолжительностью жизни у трансгенных животных с измененной активностью основных ферментативных антиоксидантов [1100]. Мыши, нокаутированные по Mn- СОД, погибали в первые 2 недели после рождения, средняя продолжительность их жизни составляла около 10 дней. У животных выявлялись тяжелые кардиомиопатии и патологии нервной системы, в их митохондриях была значительно снижена активность NADH-дегидрогеназы (комплекс I) и сукцинатдегидрогеназы (комплекс II), а также аконитазы [1136]. Другие исследователи обнаруживают только снижение активности сукцинатдегидрогеназы и аконитазы, при этом отмечают, что несмотря на повреждение митохондриальной ДНК, активность кодируемых ею белков не изменена [1035]. Имитаторы СОД и каталазы в 2—3 раза повышали продолжительность жизни мышейнокаутов по Mn-СОД [1137]. Если животные с генотипом sod2-/- погибали в первые дни после рождения, то продолжительность жизни гетерозигот была такой же, как у мышей дикого типа [1035]. Вместе с тем у мышей sod2+/- было повышено содержание 8-оксо-2'-дезоксигуанозина в структуре ядерной и митохондриальной ДНК, животные были более чувствительны к токсическому действию параквата и у них чаще развивались катаракты и лимфомы [1100]. Таким образом, хотя снижение активности Mn-СОД у гетерозиготных мышей не сказывается на продолжительности жизни, у них более выражены связанные со старением изменения.Для цитоплазматической C^Zn-СОД в разных экспериментальных системах получены неоднозначные результаты.
Эксперименты на трансгенных дрозофилах с выключенным и гиперэкспресированным геном C^Zn-СОД выявили прямой защитный эффект данного фермента. Так, продолжительность жизни нокаутированных по Cu,Zn- СОД дрозофил была снижена в 5 раз [1321], в то время как у трансгенных дрозофил с дополнительными копиями гена СОД и каталазы наблюдалось увеличение средней и максимальной продолжительности жизни [1250]. На мышах не было получено такой явной зависимости продолжительности жизни от C^Zn-СОД. Нокаутированные по C^Zn-СОД мыши имели более низкую (на 25—30 %) продолжительность жизни, дополнительное выключение кодирующего Э-СОД гена не приводило к дополнительному уменьшению продолжительности жизни двойных нокаутов [1487]. При этом для 20-месячных мышей с генотипом sod1-/- были характерны выраженная потеря мышечной массы (масса скелетных мышц задних лап снижена на 50 % по сравнению с животными дикого типа) и уменьшение двигательной активности — признаки преждевременного старения [1177]. В то же время гиперэкспрессия C^Zn-СОД у мышей не сказывалась на продолжительности жизни [781]. Некоторые исследователи считают гиперэкспрессию C^Zn-СОД причиной нейротоксичности, что в частности наблюдается у людей с синдромом Дауна (ген C^Zn-СОД человека находится в 21-й хромосоме), средняя продолжительность жизни которых снижена в 2 раза.Ни ингибирование, ни гиперпродукция экстрацеллюлярной СОД не сказывались на продолжительности жизни мышей, хотя нокауты по Э-СОД были более чувствительны к действию гипероксии (время их жизни в атмосфере чистого кислорода сокращалось на 25 % [412]), а у старых животных с гиперэкспрессией фермента наблюдалось снижение повреждений нейронов и возрастного нарушения памяти [1011]. Трансгенные мыши с избыточной экспрессией СОД, каталазы и глутатионпероксидазы обладали повышенной резистентностью к окислительному стрессу, при снижении же активности этих генов мыши имели признаки преждевременного старения. Средняя и максимальная продолжительность жизни мышей с гиперэкспрессией каталазы были повышены на 17—21 % [510].
Трансгенные мыши разных линий, в митохондриях которых был увеличен уровень каталазы, жили в среднем на 20 % дольше, чем мыши в контрольной группе, при этом у них на 25 % была снижена продукция Н2О2 и наблюдалось значительно меньше окислительных повреждений ДНК [1033, 1480]; при повышении уровня каталазы в пероксисомах или ядрах клеток эффект продления жизни был не столь выраженным и составлял 4—13 %. Гиперэкспрессия каталазы в кардиомиоцитах мышей улучшала сократительную функцию миокарда и приводила к увеличению средней продолжительности жизни на 3 месяца (около 10 %) [1787].Семейство низкомолекулярных белков тиоредоксинов служит для восстановления дисульфидных групп в белках (см. рис. 29). Мыши, нокаутированные по цитоплазматическому тиоредоксину 1 и митохондриальному тиоредоксину 2, так же как и по соответствующим тиоредоксинредуктазам, погибали в эмбриональном периоде [481, 1112, 1219]. Трансгенные мыши, несущие копии гена человеческого тиоредоксина, были более устойчивы к окислительному стрессу и имели большую продолжительность средней и максимальной жизни [1155]. Клетки их костного мозга были более устойчивы к цитотоксическому действию ультрафиолетового облучения. При этом авторы не обнаружили развития какой-либо патологии у трансгенных мышей. Мыши, нокаутированные по пероксиредоксинам, служащим для восстановления перекисей, существенно не отличаются по продолжительности жизни от мышей дикого типа, однако с возрастом у них выявляются гемолитическая анемия и злокачественные опухоли (лимфомы, саркомы, карциномы), нокауты также высокочувствительны к индуцированному липополисахаридом летальному шоку [1203, 1799].
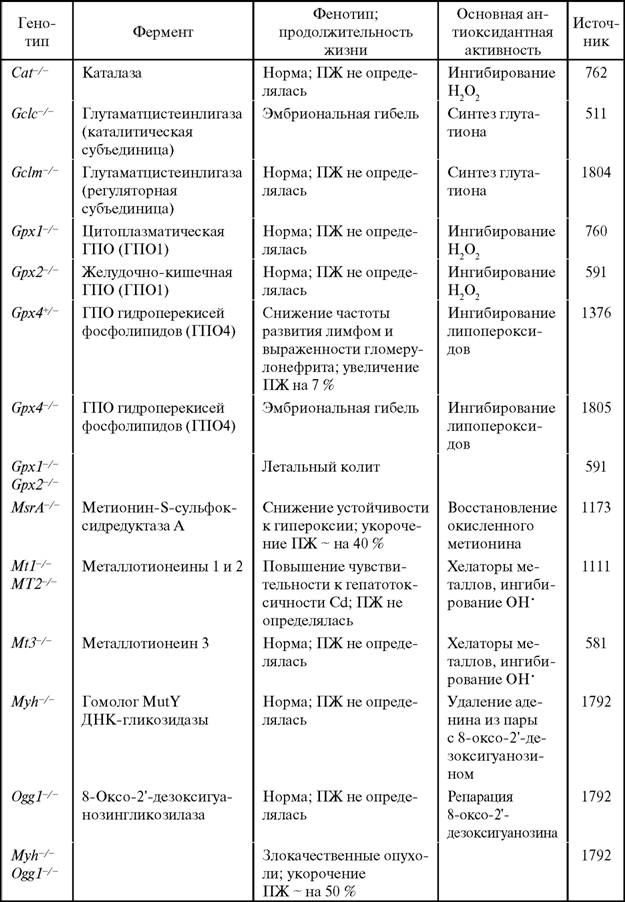
Металлотионеины играют важную роль в транспорте и обеспечении тканей ионами цинка. Хотя у нокаутированных по металлотионеинам мышей продолжительность жизни не изменена (табл. 20), возрастное снижение биодоступности ионов Zn может быть причиной усиления синтеза провоспалительных цитокинов (ИЛ-6, ФНО-α) и нарушения иммунитета [1684]. Цинк входит в состав более 300 ферментов и факторов транскрипции, важных для адаптации и развития типовых патологических процессов. Добавление сульфата цинка (22 мг/л) в питьевую воду на 50 % снижало смертность мышей от инфекций и опухолей и на 10 % повышало среднюю и максимальную продолжительности жизни [1157]. Средняя продолжительность жизни трансгенных мышей с гиперэкспрессией гена человеческого металлотионеина-IIa была на 3,65 месяца больше, чем у животных исходной линии [1803].
Антиоксидант-респонсивный элемент регулирует синтез антиоксидантов и ферментов II фазы детоксикации ксенобиотиков, он важен для поддержания внутриклеточного окислительно-восстановительного баланса [131]. Действие антиоксидант-респон-
Таблица 20
Продолжительность жизни мышей, нокаутированных по различным генам ферментов антиоксидантов и репарации