ВЫДЕЛЕНИЕ И РАЗДЕЛЕНИЕ МЕМБРАН
Выделению мембран предшествует этап разрушения клеток и тканей. Для этого подбирают методику, позволяющую не только эффективно разрушать клетки, но и сохранять нативную структуру мембран, подлежащих выделению.
При выделении большинства мембран животных клеток используют гомогенизацию в гомогенизаторах Даунса и Поттера со стеклянными стенками и тефлоновым пестиком. Для разрушения клеток применяют также вращающиеся ножевые гомогенизаторы. Предварительно свежевыделенную ткань измельчают и промывают водой. В гомогенизаторе клетки разрушаются за счет сдвиговых усилий, возникающих при продавливании суспензии через узкий зазор между тефлоновым пестиком и стеклянной стенкой. При гомогенизации следует тщательно подбирать значение pH, ионную силу и состав буферного раствора. Часто в качестве суспендирующей среды используют раствор сахарозы в концентрации 0,25 моль/л с добавлением хлорида магния, комплексообразователей (например, ЭДТА), восстановителей (дитиотреитол, р-меркаптоэтанол).С целью облегчения последующего разрушения растительных, грибных и бактериальных клеток их сначала обрабатывают ферментами, расщепляющими компоненты клеточной стенки (лизоцим, целлюлаза). Более жесткая обработка клеток предусматривает их растирание с помощью абразивных материалов (стеклянных шариков, песка, оксида алюминия), разрушение ультразвуком и путем экструзии (продавливание суспензии клеток через отверстия под давлением). Полученный гомогенат клеток процеживают и используют дальше для получения мембран.
Чаще всего для разделения мембран применяют метод центрифугирования. Мембранные частицы разделяют по скорости их седиментации (зональное центрифугирование) или по плавучей плотности (изопикническое центрифугирование). Коэффициент седиментации (S) определяется по формуле:
S = v/w2r = m(l - Vp)/f,
где v — скорость движения частицы; w — угловая скорость вращения ротора; г — расстояние до центра вращения; m — масса частицы; f — коэффициент трения; р — плотность растворителя; V — парциальный удельный объем частицы (равный увеличению объема, вызываемого прибавлением единицы массы растворяемого вещества к раствору).
Величина коэффициента седиментации зависит от нескольких факторов: концентрации раствора, скорости движения частиц, заряда, формы и массы частиц. Введено понятие стандартного значения константы седиментации, определяемое в системе, имеющей вязкость и плотность воды при +20 °С (S°o).
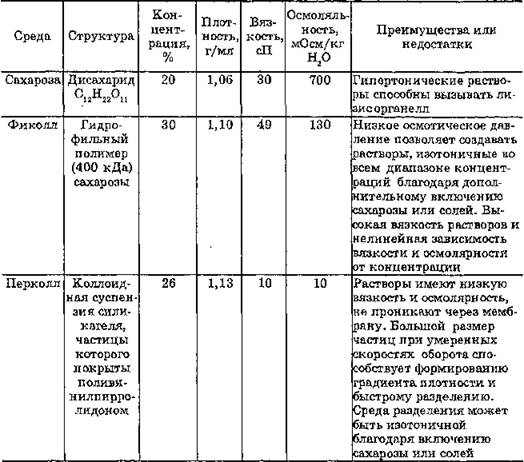
Для выделения различных мембранных структур из гомогената, имеющих разные величины коэффициента седиментации, применяют зональное центрифугирование в градиенте плотности определенных веществ. Исследуемый раствор наносят на предварительно приготовленный в центрифужной пробирке градиент плотности и центрифугируют. Градиент создается путем последовательного наслоения растворов градиентной среды уменьшающейся концентрации (плотности) в центрифужной пробирке. Субклеточные структуры разделяются на отдельные зоны в соответствии с их относительной плотностью. Для создания градиента плотности необходимо подбирать вещества в чистом состоянии, не взаимодействующие с компонентами суспензии и реагентами исследуемого раствора. Чаще всего для этого используют сахарозу, однако ее растворы с высокой концентарцией имеют большую вязкость, вследствие чего происходит дегидратация органелл или их лизис. Кроме того, серьезным недостатком этого метода является проницаемость многих органелл для сахарозы, что вызывает их осмотическое разрушение и изменение эффективной плотности. Поэтому в настоящее время для создания градиента плотности предпочитают применять другие среды: фиколл, перколл и др. (табл. 16).
Основные компоненты клетки осаждают в такой последовательности: целые клетки и их фрагменты, ядра, митохондрии, лизосомы, микротельца, микросомы (фрагменты эндоплазматической сети и плазматических мембран), отдельные типы мембранных структур.
Таблица 16
Характеристика некоторых градиентных сред для выделения мембранных компонентов методом центрифугирования
Необходимо отметить, что получаемые при разрушении клеток мембранные фрагменты способны самопроизвольно образовывать замкнутые пузырьки — везикулы.
К ним относят микросомы, субмитохондриальные частицы из внутренней митохондриальной мембраны, синаптосомы, образующиеся при отрыве нервных окончаний в области синаптических контактов. Скорость оседания таких частиц при центрифугировании определяется их размерами, зависящими от метода разрушения клеток и состава среды. Для сохранения замкнутости мембранных органелл используют среду, изоосмотичную их внутреннему содержимому (сорбитол, маннитол, сахарозу).' Следует подчеркнуть, что препаративные типы центрифуг применяют для получения и очистки клеточных органелл или макромолекул, т. е. чистых фракций (препаратов). Аналитические центрифуги позволяют анализировать распределение веществ в пробирке в течение всего опыта. Поэтому они снабжены оптической системой (пхлиреновской, интерференционной, абсорбционной).
Для выделения мембран из клеточных гомогенатов используют и другие методы: хроматографию, электрофорез, адсорбцию.
Распределительная хроматография основана на различиях в распределении разделяемых веществ между двумя фазами: подвижной (растворитель) и неподвижной (сорбент). Ее разновидностями являются хроматография на бумаге, тонкослойная, колоночная, газожидкостная. Большой разрешающей способностью характеризуется тонкослойная хроматография: она позволяет обнаружить 1 нмоль вещества. Слой сорбента (толщина до 0,5 мм) готовится из силикагеля, оксида алюминия, целлюлозы. При этом обеспечивается низкое отношение массы растворенного вещества к массе сорбента (до 1/108) и большое отношение поверхности к объему. Разрешающая способность метода колоночной хроматографии на несколько порядков ниже, чем для тонкослойной, так как отношение массы растворенных веществ к массе сорбента равно 1/50.
Адсорбционная хроматография основывается на различной способности молекул смеси адсорбироваться на поверхности носителя (силикагель, окись алюминия, активированный уголь) при пропускании через него подвижной фазы. Принцип метода ионообменной хроматографии — разновидности адсорбционной хроматографии — заключается в способности ионообменника (отрицательно заряженного катионита или положительно заряженного анионита) обратимо адсорбировать заряженные молекулы при определенных значениях pH.
Мембранные белки и углеводы разделяют на ионнообменниках на основе целлюлозы, декстрана, полиакриламида.Гелъпроникающая хроматография успешно применяется для разделения и очистки мембранных белков-ферментов, так как позволяет фракционировать вещества в широких диапазонах pH, температуры, ионной силы без адсорбции на носителе молекул разделяемых соединений. Метод основан на принципе обратного молекулярного сита: более крупные молекулы быстрее проходят через слой мелких частиц носителя, а более мелкие (с диаметром меньше или равным диаметру пор в частице) — медленнее, так как диффундируют через поры инертного материала. В качестве молекулярных сит используют декстрановые, агарозные и полиакриламидные гели. Преимущество этого метода заключается в том, что он позволяет не только разделять, но и очищать и концентрировать макромолекулы (например, при добавлении к исследуемому раствору сухих гранул геля с диаметром пор меньше, чем диаметр молекул).
Для выделения различных мембранных структур используется и аффинная хроматография. Принцип этого метода заключается в способности выделяемого вещества’ специфически связываться с лигандом, “пришитым” к нерастворимому носителю, при пропускании раствора через матрицу. В качестве последней применяют сефарозы (агарозные гели), активируемые путем связывания различных лигандов: кофакторов, ингибиторов, субстратов мембранных белков-ферментов, лектинов в случае выделения гликопротеинов; гормонов, бромциана, конканавалина А — соответственно при получении мембран, антител или целых клеток. Элюирование исследуемого вещества осуществляют в условиях диссоциации комплекса лиганд — вещество и сохранения нативной структуры выделяемого соединения.
Заряженные компоненты биомембран разделяют методом электрофореза по скорости их движения в электрическом поле, которая зависит от величины заряда, молекулярной массы и формы молекул. Максимальное разделение макромолекул (в частности, мембранных белков) обеспечивается при использовании метода диск-электрофореза в полиакриламидном геле (ПААГ).
Его название “диск” происходит от дискретного напряжения электрического поля, что обусловлено прерывистым градиентом pH в системе. Верхняя часть полимеризующегося геля в стеклянной вертикальной трубке, на которую наносится исследуемый образец, имеет меньшую концентрацию, а нижняя часть (разделяющий гель) — большую. За счет большого размера пор верхних слоев геля и большого градиента электрического поля происходит быстрое движение и накопление вещества на верхней границе разделяющего геля, в котором в зависимости от подвижности разделяемых белков образуются различные зоны, которые идентифицируют после удаления геля из трубки. Для определения молекулярной массы мембранных белков используют SDS— ПААГ-электрофорез с применением додецилсульфата натрия (ДСН, SDS), связывающегося с белком и вызывающего его диссоциацию. Подвижность белка в комплексе с ДСН будет определяться только молекулярной массой биополимера, а заряд — додецилсульфатом (при постоянстве количества связываемого любого типа белка додецилсульфатом: 1,4—1,5 г ДСН на 1 г белка). Количественные измерения проводят оптическим методом с использованием денситометра.Разновидность электрофореза — изоэлектрическое фокусирование. Метод основан на электрофоретическом разделении смеси белков в градиенте pH с достижением изоэлектрической точки (ИЭТ) каждого белка и его “фокусированием” в отдельной зоне и применяется для быстрого и эффективного фракционирования больших количеств (несколько граммов) образца на начальных стадиях его очистки.
5.3.
Еще по теме ВЫДЕЛЕНИЕ И РАЗДЕЛЕНИЕ МЕМБРАН:
- Энгельс выделял три крупных общественных разделения труда: выделение пастушеских племен
- Соединение уголовных дел, выделение уголовного дела, выделение в отдельное производство материалов уголовного дела
- Плазматическая мембрана
- Мембраны первого типа
- Мембраны второго типа
- Мембраны четвертого типа
- Проницаемость природных мембран
- Цитоскелет (мембранный каркас)
- 1.2.1. Классификация, структура и функции мембранных белков
- Мембраны третьего типа
- Часть II. Вопросы федерализма и разделения властей, затрагивающие ведомства А. Административные ведомства и разделение властей
- Роль жирных кислот мембранных липидов в холодоустойчивости растений
- Особенности структуры и функций мембранных рецепторов
- Статья 33. Выделение из унитарного предприятия
- 4.10. Стационарное мембранное фосфорил и рование
- Физико-химические и динамические свойства и функции липидов мембран
- Фоточувствителъностъ ацетилхолинэстеразы эритроцитарных мембран в присутствии фосфолипазы D и аскорбиновой кислоты
- Нарушение структурно-функциональных свойств клеточных мембран
- Синдром патологических выделений.