Физико-химические и динамические свойства и функции липидов мембран
В водной среде мембранные липиды ведут себя как анизотропные жидкости, обладающие свойствами жидких кристаллов. В жидком кристалле сочетаются особенности кристалла (дальний порядок организации, двулучепреломление) и жидкости (образование капель и текучесть).
Всем жидким кристаллам свойствен полиморфизм, т. е. они могут существовать в нескольких жидкокристаллических фазах. Даже индивидуальные очищенные липиды в гидратированном состоянии могут находиться в нескольких структурных модификациях. Преобладание того, или иного типа структуры определяется целым рядом факторов: концентрацией липида, температурой, величиной pH, ионной силой, давлением. Формирование мезоморфных структур фосфолипидов мембран зависит от соотношения липид/вода (лиотропный мезоморфизм) и от температуры (термотропный мезом-орфизм).Сочетание в молекуле липида полярного и неполярного компонентов, т. е. дифильность, обусловливает ее амфипатические свойства и, следовательно, способность к образованию мембран. Наиболее энергетически выгодным положением для молекул липидов является формирование мономолекулярного слоя на поверхности раздела масло — вода или вода — воздух (рис. 3). При достижении определенной концентрации липида — критической концентрации мицеллобразования (ККМ) его молекулы объединяются в замкнутые агрегаты — мицеллы, в которых полярные головки обращены к воде, а гидрофобные хвосты направлены внутрь. Для большинства липидов ККМ составляет менее 1 %. При более высокой концентрации формируется бимолекулярный липидный слой (ламеллярная структура). Для ламеллярной жидкокристаллической фазы (Ьц) характерно упорядоченное расположение слоистых структур при значительной неупорядоченности ацильных цепей. Считают, что именно в этой фазе находится основная масса липидов биомембран. Ламеллярная гелевая фаза (Lp) образуется при низкой температуре теми липидами, которые формируют слоистые структуры.
В этой фазе молекулы упакованы более плотно (на молекулу приходится меньшая площадь поверхности), а углеводородные цепи более упорядочены и находятся преимущественно в транс-конфигурации. Так как цепи максимально вытянуты, толщина бислоя в фазе геля выше, чем в жидкокристаллической фазе. В случае образования гексагональ-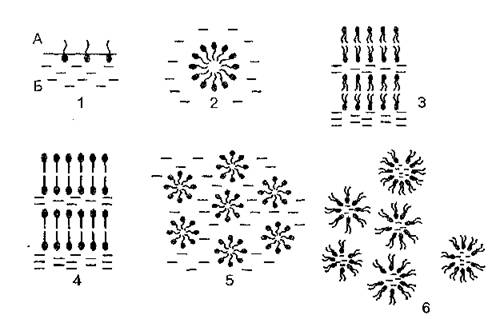
Рис. 3. Типы структурной организации водно-липидных систем: А — воздух; В — вода; 1 — монослой липидов; 2 — мицеллы фосфолипидов в воде; 3 — ламеллярная жидкокристаллическая фаза La; 4 — ламеллярная гелевая фаза Lp; 5 — гексагональная фаза типа I; 6 — гексагональная фаза типа II
ной фазы I (Hj) липидные молекулы формируют цилиндрические структуры, поверхность которых образована полярными головками и контактирует с водой. Цилиндры упаковываются с образованием гексагональной решетки. Липиды в гексагональной фазе II (Нн) также образуют цилиндры, но полярные группы обращены внутрь цилиндра и формируют водный канал. Следует отметить, что некоторые липиды (ненасыщенные фосфати- дилэтаноламины, моногалактозилдиацилглицерол) не образуют стабильные бислои, а находятся в гексагональной фазе Нп. Для изучения типов структурной организации водно-липидных систем используют метод дифракции рентгеновских лучей, а также дифференциальную сканирующую калориметрию, электронную микроскопию и метод ЯМР.
Термотропный мезоморфизм — это зависимость состояния липидных молекул от температуры. В твердой или гелеобразной фазе углеводородные цепи ориентированы строго параллельными зигзагами. После фазового перехода в жидкокристаллическое состояние, который определяется температурой и аналогичен процессу плавления, углеводородные цепи становятся подвижными. “Жидкое” состояние мембранных липидов необходи
мо для нормального функционирования всех биомембран, причем степень вязкости зависит от функциональных особенностей мембраны.
В среднем вязкость нормальной мембраны соответствует примерно вязкости оливкового масла. Термотропный ме- зоморфизм существенно зависит от природы жирных кислот и полярной головки липидов. Так, увеличение числа двойных связей и укорочение углеводородных цепей приводят к снижению температуры фазового перехода.Вышеописанные свойства липидов взаимосвязаны друг с другом. Температура фазового перехода зависит от содержания воды в анализируемой системе.
Различные липиды способны к формированию разных мезоморфных структур, что обусловлено особенностями строения молекул и соотношения объемов полярных головок и углеводородных хвостов. Липиды с электронейтральной головкой (фосфатидилхолин, фосфатидилэтаноламин, сфингомиелин) образуют ламеллярную фазу. Липиды с отрицательно заряженными головками вследствие действия электростатических сил отталкивания формируют мицеллярные или гексагональные структуры. В случае равенства объемов, занимаемых полярными головками и углеводородными хвостами, молекулы липида имеют цилиндрическую форму и образуют бислой (фосфатидилхолин). Если объем полярной головки больше объема углеводородных цепей (лизо- фосфолипиды), то молекула имеет форму перевернутого конуса и в водном растворе находится в мицеллярной фазе. Если объем полярной головки меньше объема углеводородных цепей (ненасыщенный фосфатидилэтаноламин, кардиолипин в присутствии ионов Са2+, фосфатидная кислота), то молекула липида имеет форму конуса и образует гексагональную фазу типа II. В целом способы упаковки различных липидов с учетом геометрической формы их молекулы определяются следующими параметрами: молекулярным объемом неполярной части молекулы V, максимальной длиной этого участка 1, оптимальной площадью поверхности, занимаемой полярной головкой So. Критический параметр упаковки липидов представляет собой величину V/1SO.
Фазовые переходы липидов сопровождаются значительным повышением ионной проницаемости мембран. По-видимому, мицеллярная и гексагональная фазы, способные формировать сквозные поры, более проницаемы для ионов и воды, чем бислойная ламеллярная структура.
Способность мембранных фосфолипидов к образованию в водной среде мезоморфных структур в физиологическом оптимуме температур обусловливает ион-регуляторную функцию мембранных липидов. Так, для заряженных фосфолипидов, фосфатидилэтаноламина и лизолецитина возможны фазовые переходы из бислоя в мицеллярную и гексагональную фазы. Для фосфатидилхолина характерен вид бислойной ламеллы с фазовыми переходами термотропного характера из жидкокристаллического состояния в гель и обратно. В связи с этим выяснение молекулярных механизмов полиморфных переходов в липидной фазе необходимо для изучения процессов транспорта веществ через биомембраны.Динамическое состояние липидного бислоя, являющееся основой функционирования мембраны, определяется целым рядом факторов: вращательной и латеральной диффузией отдельных молекул фосфолипидов, подвижностью их углеводородных цепей, транс-гош-изомеризацией остатков жирных кислот. Лабильность мембранных белков, в свою очередь, зависит от фазового состояния и вязкости липидного матрикса мембраны. С помощью метода ЭПР показано, что для молекул фосфолипидов в мембранах характерны движения двух типов:
1) латеральная диффузия — перемещение в пределах одного слоя липидной фазы параллельно поверхности мембраны;
2) трансбислойный переход типа “флип-флоп” — перемещение из одного монослоя липидного матрикса в другой. Его частота в везикулах яичного лецитина около 210“6 с'1, он происходит на 12 порядков медленнее, чем латеральная диффузия. “флиц-флоц”-переходы молекул фосфолипидов характеризуются высокими значениями энергии активации (= 80 кДж/моль), а обмен в пределах одного монослоя липидов — более низкими (—21 кДж/моль).
Движение углеводородных цепей состоит из торсионных (вращательных, крутильных) колебаний с относительно малой амплитудой (< 20°) вокруг каждой из связей (время корреляции вращения slO'14 с) и транс-гош-изомеризации отдельных звеньев (s 1О“10 с). Минимальной энергией обладает транс-, а максимальной — цис-конформация углеводородных цепей.
Углеводородные цепи в полной транс-конформации представляют собой линейные структуры. Гош-конформации (гош “+” и гоні поворот на ±120° относительно транс-конформации) мало превышают по уровню энергии транс-конформацию (на 2—3 кДж/моль), но эти состояния разделяет энергетический барьер высотой примерно 12—17 кДж. Появление одиночной гош-конформации приводит к искривлению пространственной конформации углеводородной цепи примерно на 120°, что затрудняет появление одиночных гош-конформаций. При последовательном повороте цепи на +120 и -120°, т. е. при образовании двух гош-конформаций (“+” и пространственная структура цепи сохраняется прямолинейной. Участок цепи, находящийся в гош “+”- транс-гош “-’’-конформации, формирует в углеводородной цепи петлю, называемую кинком. Образование кинка сопровождается уменьшением эффективной длины углеводородной цепи примерно на 0,127 нм, а объем, занимаемый молекулой липида, увеличивается (рис. 4). Формирование кинков — кооперативный процесс, приводящий к возрастанию разупорядоченности углеводородной зоны и ее плавлению. Этот процесс стимулируют двойные (цис-) связи в ненасыщенных цепях мембран. Для образования кинка в таких участках необходимо появление лишь одной гош-конформации при искривлении цепи на 80". Поэтому высокая текучесть углеводородной зоны мембран в жидкокристаллическом состоянии объясняется возрастанием амплитуды крутильных осцилляций вокруг С—С-связей, появлением гош-конформаций.С текучестью мембран тесно связаны функциональная активность мембраносвязанных ферментов, а также функционирование систем пассивного транспорта. Формирование кинков, обусловленное появлением в липидном бислое флуктуирующих объемов, является одним из возможных механизмов трансмембран-
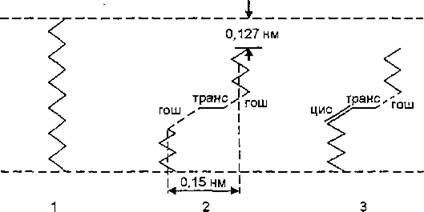
Рис. 4. Конформация углеводородных цепей липидов в мембране: 1— полная транс-конформация; 2— гош-транс-гош-конформация; 3 — цис-транс-гош-конформация
ного переноса воды и ионов.
Вопросы, касающиеся выяснения роли мембранных липидов в процессах транспорта ионов и механизмов образования ионных каналов при фазовых превращениях липидов, подробно изложены в монографии В. Ф. Антонова “Липиды и ионная проницаемость мембран” (1982).Динамическая модель липидного бислоя показана на рис. 5. Липидный матрикс состоит из области полярных групп Р и области углеводородных цепей Н. Углеводородная часть представлена как бы тремя слоями: двумя упорядоченными толщиной - 0,8 нм, прилегающими к полярной области, и центральным “изотропным”, подобным короткоцепочечному жидкому углеводороду. Толщина этого центрального слоя для бислоев дипальмитоил-, димиристоил- и дилауроиллецитина составляет соответственно 0,85 и 0,2 нм. По- видимому, существование “изотропной” области необходимо для того, чтобы бислой был непроницаем для ионов и малых молекул. Повышение проницаемости липидного матрикса связано с образованием кластеров и динамических дефектов. Кластеры — это области с сохраняющимся ближним порядком молекул, упаковка которых близка к кристаллической. Кластеры представляют собой динамические (мгновенные) образования с временем жизни - 10'7 с, включающие 40—60 углеводородных цепей (20—30 молекул фосфолипидов). В них ближайшие к полярным головкам участки углеводородных цепей имеют несколько более плотную упаковку, чем в “твердом” бислое. По направлению к центру бислоя плотность упаковки уменьшается, как и на границах кластеров, кото-
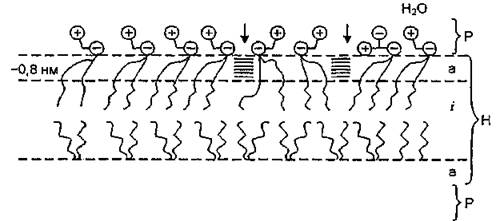
Рис, 5. Динамическая организация липидного бислоя: Р — области полярных групп; Н — области углеводородных цепей; а — упорядоченная “анизотропная” область; і — “изотропная” область; -I — динамические дефекты
рые непрерывно распадаются и образуются, перемещаясь вдоль бислоя. При этом граничные области между кластерами представляют собой динамические дефекты в бислое. Последние также носят динамический характер, поэтому степень проницаемости липидной мембраны должна зависеть от времени жизни дефектов. Увеличение времени их жизни путем фиксации части кластеров при отвердевании бислоя приводит к резкому (на 1—2 порядка) возрастанию проницаемости мембраны.
Необходимо отметить, что на ориентацию и динамику полярных головок липидов влияет образование межмолекулярных водородных связей на поверхности мембраны. Донорами и акцепторами при образовании этих связей могут служить фосфати- дилсерин, фосфатидилэтаноламин, гликолипиды. Однако не ясно, каким образом водородные связи воздействуют на структуру мембраны. С точки зрения термодинамики, основной силой, стабилизирующей гидратированные липидные агрегаты, являются гидрофобные взаимодействия. К другим стабилизирующим факторам относятся водородные связи и ван-дер-ваальсовы силы (короткодействующие слабые силы притяжения между соседними гидрофобными цепями).
Важнейшим свойством липидного бислоя мембран является структурная асимметрия — различное распределение липидов между внутренним и наружным монослоями. Анализ распределения фосфолипидов в мембранах микросом, аппарата Гольджи, лизосом, ядер, митохондрий показывает, что фосфатидилэтаноламин и фосфатидилсерин расположены преимущественно на цитоплазматической стороне, сфингомиелин, кардиолипин и фосфатидилинозитол — на внутренней, топологичной наружной стороне плазматических мембран, а фосфатидилхолин более или менее равномерно распределен между обеими сторонами. В наружном монослое липидов мембран эритроцитов человека содержится 44 % фосфатидилхолина, 44 % сфингомиелина и 12 % фосфатидилэтаноламина, во внутреннем — 48 % фосфатидилэтаноламина, 28 % фосфатидилсерина, 10 % сфингомиелина и 14 % фосфатидилхолина.
Углеводородные цепи, входящие в состав фосфатидилхолина и сфингомиелина, более насыщены по сравнению с теми, которые находятся в составе фосфатидилэтаноламина и фосфатидилсерина. В связи с этим асимметрия в распределении полярных головок сопровождается асимметрией распределения жирнокислотных хвостов. Это может привести к тому, что текучесть внутреннего монослоя будет несколько больше, чем внешнего. Известно также, что отрицательно заряженный фосфатидилсерин локализован во внутреннем монослое, следовательно, две стороны бислоя существенно различаются и по заряду. Феномен асимметричного распределения липидов необходим для проявления функциональной активности многих мембраносвязанных ферментов. Так, при активации протеинкиназы С она связывается с цитоплазматической стороной плазматической мембраны, где концентрируется фосфатидилсерин, необходимый для работы фермента. Асимметрия распределения липидов определяется не только ли- пид-белковым взаимодействием, но и асимметрией липидного синтеза, различиями ионного состава вне- и внутриклеточной среды. Асимметрия бислоя обеспечивается ферментами липидного обмена и липидпереносящими белками (липазами, системами обмена холестерина, метилазами фосфатидилэтаноламина).
Липиды в биомембранах выполняют множество функций. Во- первых, они обеспечивают структурную организацию и стабильность клеточных мембран. Во-вторых, выполняют барьерную и транспортную функции. В-третьих, играют фундаментальную роль в передаче информации и регулировании метаболических процессов в клетке. Последняя функция мембранных липидов включает участие их в реакциях биосинтеза; поддержании оптимальной активности белков-ферментов мембран; выполнении рецепторных функций, обеспечивающих проявление иммунологических свойств и ответственных за взаимодействие клеток; а также в процессах накопления, передачи и хранения энергии. Липиды участвуют в механизмах кратковременной и долговременной памяти. В дальнейшем вопрос о выполнении липидами регуляторной роли в различных процессах метаболизма будет рассмотрен более подробно в главах 2, 3.
1.2.