4.10. Стационарное мембранное фосфорил и рование
До сих пор речь шла о механизме «элементарного акта» синтеза АТР Величина трансмембранного Ддн+ не имеет никакого отношения к элементарному акту. Этот градиент непрерывно поставляет ионы водорода для повторяющихся «ударов», реализуя, таким образом, возникновение неравновесных состояний и их последующую конформационную релаксацию.
В монографии [31] была предложена следующая упрощенная схема циклической работы АТРсинтазы (рис. 4.28). Эта схема объясняет
многократное образование АТР после создания разницы pH по обе стороны мембраны везикул, содержащих факторы F,, доступные внешней водной среде. Кстати, градиент Ддн в некоторых условиях включает
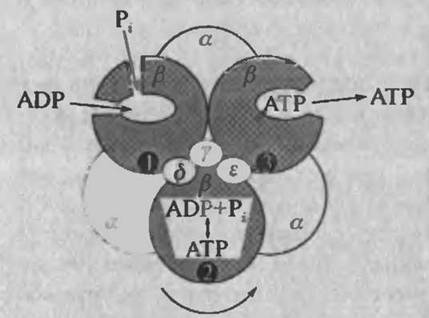
Рис. 4 28. Схема циклического функционирования АТРсинтазы
не только ДрН, но и ірадиент трансмембранного электрического потенциала Д0. Анализ приводит в этом случае к тому же выводу. Д зависит от распределения зарядов вблизи активного центра и определяет величину рК кислотных групп. Пересечение уровня pH при быстром уменьшении рК во время конформационной релаксации приведет к тому же эффекту, что и пересечение уровня рК при быстром повышении pH
На схеме рис. 4.28 мы рассмотрели только один центр, связывающий АТР и ADP с двумя протон акцепторными группами А~ и В . Присоединение ADP и выброс АТР в водную фазу играет роль «переключателя», который попеременно осуществляет контакт F, то с внутренней (кислой), то с внешней («щелочной») водными фазами Различные положения переключателя символизируют возможность протонных групп А и В контактировать с внутренней и с внешней средой. Это показано на рис 4.27 в виде двух клапанов anb. Клапан а осуществляет контакт функциональной кислотной группы с внутренним объемом везикулы, а b — с внешней средой. Открытое положение а с закрытым положением Ь (состояние 1) указывают, что средняя скорость протонирования А " и В’ из внутреннего пула протонов согласуются с соответствующими скоростями диссоциации протонов из АН и ВН во внешнюю среду Перекрытие а может быть вызвано присоединением субстратов фосфорилирования, в то время как их удаление из F, обеспечивает контакт А' и В“ с внутренней областью везикулы (клапан а открыт).
Хорошо известно, что при любых условиях, неблагоприятных для синтеза АТР (например, отсутствие Pt),связывание АТР или ADP с активным центром Ft хлоропластов блокирует выход протонов из тилакоида через канал Fo [71,72].
Рассмотрим теперь последовательность событий во время функционирования АТРсинтазы, связанной с мембраной, при наличии трансмембранной разности pH. Положение 1 на рис. 4.27 соответствует квазиравновесному состоянию фермента: нет субстратов фосфорилирования в активном центре, функциональные кислотные группы протонированы, благодаря их контакту с «кислым» внутренним объемом везикулы (pH,- < рКа), а открыт. Быстрая утечка протонов во внешнюю водную фазу через АТРсинтазу предотвращается барьером, запрещающим контакт АН группы с внешней средой (клапан b закрыт). Присоединение субстратов фосфорилирования к активному центру Fj (переход 1 —► 2) делает возможными два события:
1. Бесплатное превращение ADP и Р, в прочно связанную АТР
(переход 2 —> 3).
2. Блокирование пути протона через а (клапан а закрыт).
Таким образом, после связывания субстратов группа АН диссоциирует, и становится возможным выход протонов в «щелочную» водную фазу (рКа < рН0, клапан b открыт — переход 3 —► 4) в соответствии с хорошо известными экспериментальными фактами [71,72]: процесс фосфорилирования ADP ускоряет выход протонов через Fo. Эта модель предполагает, что связывание субстратов стимулирует выход протонов во внешнюю среду. В неравновесном состоянии 4 фермент содержит прочно связанную молекулу АТР. Состояние 4 энергетически невыгодно из-за избытка отрицательных зарядов в белковой глобуле. В ходе последующей релаксации фермента молекула АТР отщепляется от F, во внешнюю водную фазу. Таким образом, фермент достигает состояния 5: а открыт, b закрыт. После последующего протонирования кислотных групп (переход 5 —> 1) цикл будет повторяться, пока выполняются условия: pH, < рКа < рН0.
Попробуем теперь ответить на важный, хотя скорее риторический вопрос: что является движущей силой образования АТР в ходе мембранного фосфорилирования? Обычный ответ в почти любой публикации, касающейся этой проблемы, таков: ДрН (Д^н+) между двумя сторонами мембраны. Этот ответ ошибочен. Разность концентраций и кинетической энергии протонов являются тепловыми составляющими полной внутренней энергии системы (терминология МакКлэйра) и не могут быть использованы для совершения работы. Должно присутствовать механическое устройство для преобразования теплового компонента в сохраняемый. Повторим еще раз, что на молекулярном уровне механические и электрические силы неразличимы [32]. В настоящее время конструкция и способ работы этого устройства известны.
4.10.