Фоточувствителъностъ ацетилхолинэстеразы эритроцитарных мембран в присутствии фосфолипазы D и аскорбиновой кислоты
К группе естественных модификаторов, изменяющих липидный состав мембран в нативной клетке, относятся липидперено- сящие белки, ферменты обмена фосфолипидов — фосфолипазы, диметилазы и др., а также системы обмена холестерина.
Вследствие их деятельности осуществляются выраженное изменение содержания лизоформ отдельных липидов, накопление в бислое жирных кислот, обладающих детергентным действием, изменение соотношения фосфатидилхолин/фосфатидилэтаноламин и фосфолипиды/холестерин. Все эти факторы управляют микровязкостью мембраны и оказывают влияние на подвижность ее компонентов.Фосфолипазы — обширный класс липолитических ферментов, имеющих первостепенное значение для регулирования разнообразных процессов жизнедеятельности всех живых организмов. Это связано с многообразием их функций. Во-первых, они участвуют в обновлении мембранных фосфолипидов, что определяет стабильность и биохимическую активность мембран и в конечном итоге функциональное состояние целой клетки. Во-вторых, продукты фосфолипазной реакции (жирные кислоты, лизо- фосфатидилхолин, холин, диглицерид, фосфорилхолин и др.) являются мощными эффекторами мембранных процессов. В-третьих, фосфолипазам принадлежит ключевая роль в биосинтезе простагландинов, лейкотриенов и других продуктов превращения арахидоновой кислоты. Так, реакцию гидролиза фосфолипидов, приводящую к образованию свободной арахидоновой кислоты, катализирует фосфолипаза А2, Эта реакция является лимитирующей стадией в “каскаде” ферментативных реакций биосинтеза физиологически активных эйкозаноидов.
Вместе с тем всестороннее изучение структурно-функциональных свойств фосфолипаз имеет важное не только теоретическое, но и практическое значение для медицины и фармакологии. Многие патологические процессы, в том числе ишемия и воспаление, сопровождаются увеличением активности внутриклеточных фосфолипаз.
Предполагают, что продукты метаболизма арахидоновой кислоты участвуют в защитной реакции организма при облучении, а также комбинированном радиационно-термическом поражении его отдельных систем.Фосфолипазы (фосфатидацилгидролазы, КФ 3.1) — ферменты, катализирующие гидролитическое расщепление сложноэфирных связей в различных фосфолипидах. Фосфолипазы Ар А2, В, С, D различают в зависимости от того, какая из четырех эфирных связей, имеющихся в молекулах фосфатидов, гидролизуется ферментом. Существует также цифровая номенклатура фосфолипаз соответственно положению расщепляемой сложноэфирной связи: 1, 2, 3, 4 (рис. 39).
Фосфолипазы различного происхождения способны также катализировать реакции ферментативного трансалкилирования (фосфолипаза D) и трансацилирования (другие фосфолипазы).
Липолитические ферменты гидролизуют водонерастворимые субстраты, следовательно, они должны функционировать на границе раздела фаз. Обычно их молекулы состоят из трех участков: гидрофобного, который создает необходимую ориентацию фермента на поверхности субстрата, и активного центра, находя-
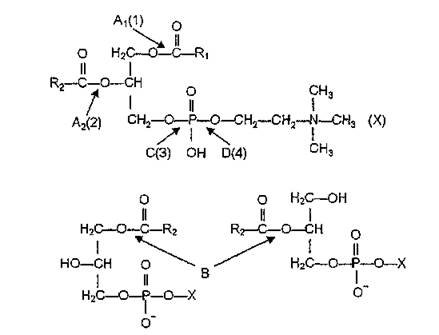
Рис. 39. Схема действия и номенклатура фосфолипаз
щегося вблизи головки, но не идентичного ей, и гидрофильного хвоста, стабилизирующего ориентацию белковых молекул.
По аналогии с другими гидролитическими ферментами для фосфолипаз предполагают наличие различных типов активных центров: серингистидинового, карбоксил-карбоксилатного, Zn2'-Kap6o- ксилатного, цистеин-гистидинового. Именно с участием этих групп происходит осуществление каталитического акта гидролитическими ферментами.
Фосфолипаза D (фосфатидилхолин-фосфатидатгидролаза, фосфолипаза 4) гидролизует эфирную связь между фосфатной группой и гидрофильным спиртом в фосфоглицеридах и соответствующую связь в сфингомиелинах. Фермент был впервые обнаружен в корнеплоде моркови и листьях шпината, он содержится также и в других растениях: свекле, брюссельской и савойской капусте.
Субстратами фосфолипазы D являются фосфолипиды и их лизопроизводные. Легкость гидролиза уменьшается в последовательности: фосфатидилглицерин, лизофосфатидилхолин, дифос- фатидилглицерин. Ферментативные процессы с участием растворимых фосфолипаз и нерастворимых субстратов протекают на границе раздела фаз: липид — вода. Эти процессы предполагают первоначально реакцию иммобилизации водорастворимого фермента на поверхности субстрата. Поэтому фосфолипаза D может использоваться в качестве модели для исследования гетерогенного ферментативного катализа и изучения нримембранных ферментов.Рис. 40. Ферментативная активность ацетилхолинэстеразы эритроцитарных мембран, модифицированных фосфолипазой D, после УФ- облучения в дозе 1,5 кДж/м2: 1 — контроль (нативный фермент); 2 — УФ-облучение; 3 — модификация фосфолипазой D; 4 — УФ-облуче- ние модифицированных фосфолипазой D мембран
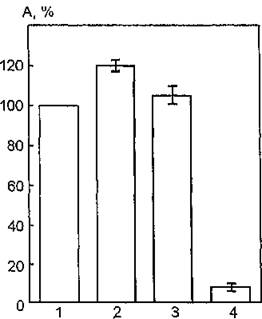
На рис. 49 показаны УФ-индуцированные изменения функциональной активности АХЭ мембран эритроцитов, модифицированных фосфолипазой D, в интактном состоянии и после УФ- облучения в дозе 1,5 кДж/м2. Предварительная обработка эритроцитарных мембран раствором фосфолипазы D в концентрации 10*fi моль/л (pH 5,6; 0,03 моль/л СаС12) с последующим удалением модифицирующего -агента путем центрифугирования практически не влияет на величину каталитической активности мембранной АХЭ: уровень изучаемого параметра — 105 % по отношению к таковому для нативных мембран (100 %). Однако УФ-облучение эритроцитарных мембран, обработанных фосфолипазой D, индуцирует резкое снижение функциональной активности АХЭ до 8 %. Следовательно, воздействие УФ-света на мембраносвязанный фермент после модификации липидной фазы мембран вызывает его инактивацию, т. е. выявляется эффект, противоположный наблюдаемым изменениям (активация) исследуемого параметра при облучении интактных мембран.
Таким образом, в случае химической модификации липидного компонента мембраны путем гидролиза фосфоглицеридов и сфингомиели- на как на внешней, так и на внутренней поверхности мембраны фоточувствительность эритроцитарной АХЭ существенно изменяется.Аналогичные результаты были получены и при исследовании функциональной активности АХЭ мембран эритроцитов, модифицированных фосфолипазой, в нативном состоянии и после УФ-облучения в дозе 3,0 кДж/м2 (рис. 41). Воздействие УФ-излучения в дозе 3,0 кДж/м2 на эритроцитарные мембраны, обработанные фосфолипазой D, приводит к практически полному ингибированию фермента. Следовательно, и в этом случае модификация липидной фазы мембран вызывает инактивацию АХЭ — эффект, противоположный изменениям активности белка, регистрируемым при облучении интактных мембран.
На рис. 42 показаны изменения ферментативной активности мембраносвязанной АХЭ после обработки мембран фосфолипазой D и воздействия УФ-света в дозе 4,5 кДж/м2. Видно, что в результате УФ-облучения модифицированных мембран фоточувствительность фермента изменяется незначительно. Активность АХЭ в интактных мембранах после облучения снижается на 44 %, а в обработанных фосфолипазой — на 52 % по отношению к контрольному образцу.
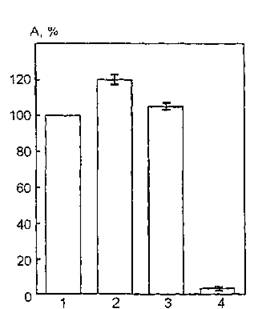
Рис. 41. Каталитическая активность ацетилхолинэстеразы эритроцитарных мембран, модифицированных фосфолипазой D, после УФ- облучения в дозе 3,0 кДж/м2: 1 — контроль (нативный фермент); 2— УФ-облучение; 3 — модификация фосфолипазой D; 4 — УФ-облучение модифицированных фосфолипазой D мембран
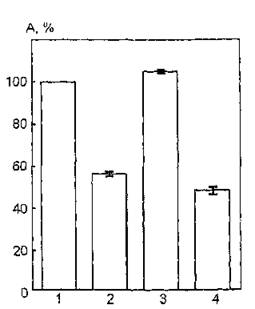
Рис. 42, Функциональная активность ацетилхолинэстеразы эритроцитарных мембран, модифицированных фосфолипазой D, после УФ-облучения в дозе 4,5 кДж/м2: 1 — контроль (нативный фермент); 2 — УФ-облучение; 3 — модификация фосфолипазой JD; 4 — УФ-облучение модифицированных фосфолипазой D мембран
Рис.
43. Фоточувствительность ацетилхолинэстеразы интактных и модифицированных фосфолипазой D эритроцитарных мембран: 1 — интактные мембраны; 2 — модифицированные мембраны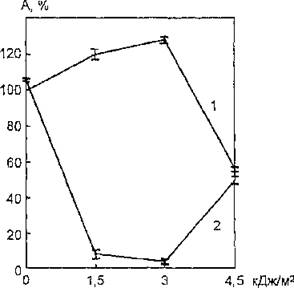
На рис. 43 представлены данные, характеризующие зависимость функциональных свойств мембранной АХЭ от дозы облучения для интактных и обработанных фосфолипазой D эритроцитарных мембран. Полученные результаты свидетельствуют о том, что УФ-чувствительность мембраносвязанной ацетилхолинэстеразы существенным образом зависит от структурного состояния фосфолипидных компонентов биомембраны.
Фосфолипаза D катализирует отщепление гидрофильного спирта от фосфоглицеридов и сфингомиелинов. В результате отщепления полярных головок от молекул фосфолипидов могут нарушаться электростатические взаимодействия фермента с молекулами окружающих его фосфолипидов, в частности, кислого липида — фосфатидилсерина, являющегося, по-видимому, аннулярным липидом для ацетилхолинэстеразы. Фосфатидилсерин, а также фосфатидилэтаноламин локализованы преимущественно во внутренней половине липидного бислоя. Можно предположить, что гидролиз фосфолипидов и фосфатидилсерина не вызывает конформационных изменений молекул фермента, затрагивающих его активный центр, поэтому активность АХЭ при обработке мембран фосфолипазой практически не изменяется. Необходимо отметить, что обработка мембран фосфолипазой индуцирует изменения упаковки и подвижности фосфолипидов, вязкости и асимметрии липидной фазы, белок-липидных взаимодействий. Воздействие УФ-излучения на модифицированные мембраны приводит к нарушениям в функционировании мембраносвязанной АХЭ, отличающимся по направленности от таковых при облучении интактных мембран. Эти нарушения являются результатом изменения конформационного состояния продуктов гидролиза фосфолипидов в мембране при воздействии УФ-света.
Е. А. Лапшина, И. Б. Заводник (1995) установили, что взаимодействие свободных жирных кислот (пальмитиновой, лаури- новой, каприловой) и их производных с эритроцитарной мембраной вызывает ингибирование АХЭ цельных эритроцитов и изолированных эритроцитарных мембран.
Авторы считают, что воздействие жирных кислот и их производных на мембрану приводит к изменению белок-липидных контактов, возмущению нативного липидного окружения фермента, нарушению максимальной комплементарности между гидрофобной поверхностью мембранных белков и их липидным окружением. Обработка эритро-цитов малоновым диальдегидом, способным модифицировать амино- и сульфгидрильные группы белков, образуя ряд стабильных и нестабильных аддуктов, внутри- и межбелковые сшивки, приводила к ингибированию активности АХЭ по бесконкурентному механизму.
Таким образом, выявляется различная УФ-чувствительность АХЭ интактных и модифицированных фосфолипазой D эритроцитарных мембран, обусловленная нарушением нативной конформации фермента вследствие воздействия на него фотохимических продуктов гидролиза фосфолипидов.
Большой интерес в настоящее время представляют вопросы, касающиеся всестороннего изучения процессов фотомодификации отдельных компонентов биомембран в присутствии химических соединений с различным механизмом действия, способных инициировать, усиливать или ослаблять фотохимические реакции. Одним из факторов системы низкомолекулярных регуляторов, выполняющих роль инициаторов, катализаторов, ингибиторов и влияющих на стадию инициирования, разветвления и обрыва цепи свободнорадикальных реакций, являются низкомолекулярные компоненты антиоксидантной системы (см. раздел 3.3).
Содержание аскорбиновой кислоты, обладающей чрезвычайно широким спектром антиоксидантных свойств, составляет в плазме крови 50—200 мкмоль/л. Вместе с тем известно, что низкомолекулярные антиоксиданты способны проявлять свои защитные
Рис. 44. Ферментативная активность ацетилхолинэстеразы эритроцитарных мембран в присутствии аскорбиновой кислоты в различных концентрациях (моль/л): 1 — контроль (нативные мембраны); 2 — 0,5-Ю’3; 3 — 10~4; 4 — 10~5; 5— 10-°; 6— 10"й
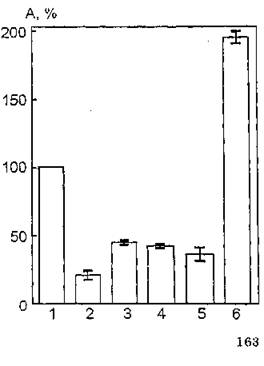
функции в широком интервале концентраций: от 10~2 до 10“18 моль/л. На рис. 44 показаны изменения функциональной активности АХЭ эритроцитарных мембран в присутствии аскорбата в различных концентрациях. Из анализа данных, представленных на этом рисунке, следует, что добавление к суспензии эритроцитарных мембран аскорбиновой кислоты в концентрациях 0,5-ПУ'1, IO-1*, 10"s, 10"а моль/л индуцирует статистически достоверное снижение уровня каталитической активности АХЭ. Использование экзогенного агента в минимальной концентрации (10~8 моль/л) приводит к активации фермента на 93 % по отношению к контрольному образцу (100 %).
Витамины А,С, D и Р, при окислении и аутоокислении которых образуются промежуточные радикальные формы (например, пероксид водорода), могут выполнять роль инициаторов окисления и ускорять ПОЛ, увеличивая скорость зарождения цепей в мембранах. Кроме того, в присутствии ионов железа и меди, содержащихся в эритроцитарной мембране, аскорбиновая кислота становится мощным прооксидантом, что указывает на необходимость in vivo надежной секвестрации свободных ионов металлов переменной валентности. Проявление аскорбатом анти- и про- оксидантных свойств зависит также от концентрации субстрата и условий протекания окислительных реакций.
Следовательно, в интервале используемых концентраций 0,5'10-3—10"8 моль/л аскорбиновая кислота проявляет проокси- дантные свойства, что находит отражение в снижении функциональной активности мембраносвязанной ацетилхолинэстеразы. Промежуточные радикальные формы, образующиеся при окислении аскорбата, усиливают и ускоряют ПОЛ, в результате которого накапливаются пероксидные продукты. Ацетилхолинэстераза принадлежит к числу ферментов мембран, легко инактивируемых при пероксидном окислении ненасыщенных жирных кислот. Наиболее важные изменения в белковых молекулах, вызываемые окисленными липидами, заключаются в образовании комплекса окисленный липид — белок, ассоциации белковых молекул и разрушении аминокислот, в частности, содержащих SH-группы.
На рис. 45 представлены данные, полученные при исследовании уровня функциональной активности АХЭ эритроцитарных мембран, УФ-облученных в дозе 4,5 кДж/м2 в присутствии аскорбиновой кислоты. УФ-облучение интактных мембран вызывает снижение активности фермента на 44 %. Совместное дей-
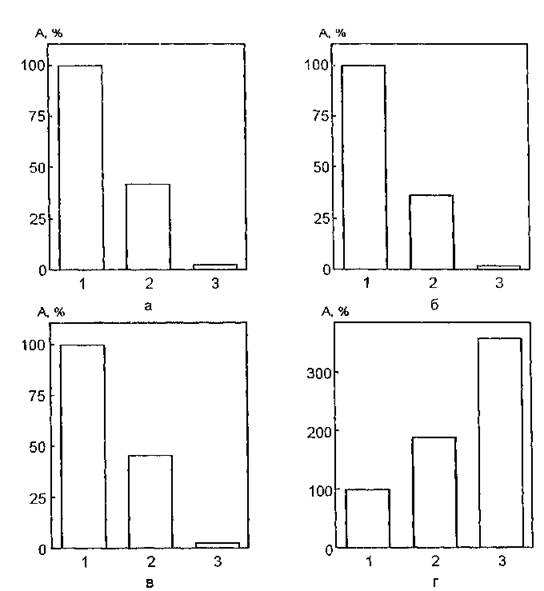
Рис. 45. Изменения каталитической активности мембраносвязанной ацетилхолинэстеразы, УФ-облученной в присутствии аскорбата: 1 — контроль (нативные мембраны); 2— в присутствии аскорбата; 3 — УФ-об- лучение в присутствии аскорбата. Концентрация аскорбата, моль/л: а — 10~4; б — Ю'5; е— 10-“; г — Ю'8
ствие УФ-излучения и аскорбата в концентрациях 10"4, 10~в, 10'° моль/л индуцирует полное ингибирование мембранной АХЭ. УФ-облучение мембран эритроцитов в комплексе с модифицирующим агентом (10"8 моль/л) приводит к резкому повышению функциональной активности белковой молекулы.
При изучении ПОЛ в мембранах митохондрий и микросом было установлено, что существенное усиление накопления перок-
сидных продуктов может быть вызвано добавлением солей двухвалентного железа, аскорбиновой кислоты и соединений, содержащих сульфгидрильные группы, например, цистеина или глутатиона. В 1959 г. Оттоленги обнаружил, что накопление пероксидов связано с наличием в изучаемой среде двухвалентного железа. Он предположил, что ионы Fe2+ оказывают каталитическое действие на образование пероксидов. При этом Fe2+ окисляется до Fe3+, а функция аскорбиновой кислоты заключается в регенерации ионов за счет обратного восстановления Fe;,+ до Fe2+. Ведущую роль ионов Fe2+ в процессе ПОЛ в биомембранах определяет совокупность следующих реакций:
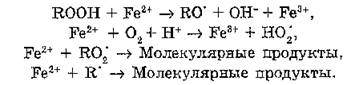
Следует подчеркнуть, что скорость ПОЛ определяется соотношением концентраций ионов Fe2+, гидропероксидов, свободных радикалов RO2‘. Ионы железа играют одновременно функцию про- и антиоксидантов. Их антиокислительное действие проявляется только при достаточно высоких концентрациях Fe2+ (>10‘5—10"4 моль/л).
По всей вероятности, ингибирование мембраносвязанной АХЭ при ее УФ-облучении в присутствии аскорбата (И)"4—10"® моль/л) представляет собой результат параллельного протекания пероксидного фотоокисления ненасыщенных жирных кислот фосфолипидов и аскорбатзависимого ПОЛ с участием ионов Fe2+. Причем эти процессы взаимоусиливают друг друга. Антиоксидантный и активирующий эффекты аскорбата по отношению к уровню функциональной активности мембраносвязанной АХЭ зарегистрированы только в присутствии экзогенного агента в концентрации 10~8 моль/л. В данном случае реализуются процессы обезвреживания аскорбиновой кислотой активных форм кислорода (супероксидного анион-радикала, синглетного кислорода, гидроперок- сидного радикала, гидроксильного радикала, пероксидных радикалов липидов), восстановления а-токоферильного радикала.
Изучение фоточувствительности мембраносвязанной АХЭ в присутствии химических агентов, модифицирующих липидную фазу мембраны, позволяет расширить современные представления о молекулярных механизмах регулирования активности важнейших компонентов биомембран. На основании собственных эк- 166
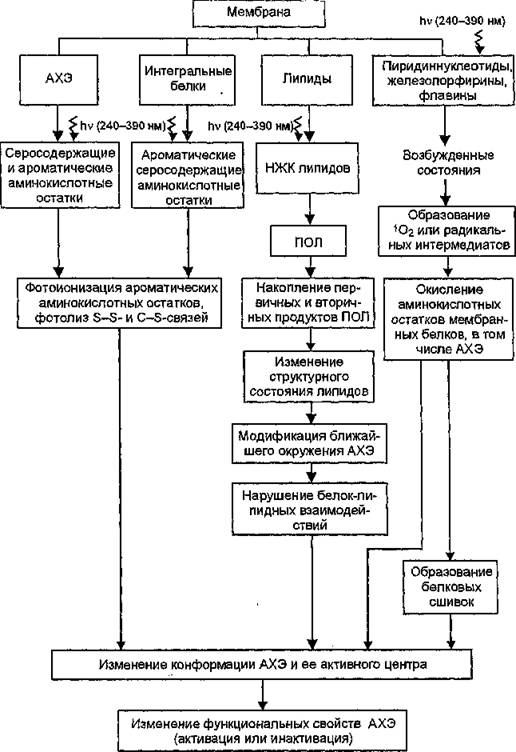
Рис. 46. Схема процессов, приводящих к фотомодификации мембранной ацетилхолинэстеразы
спериментальных и литературных данных предложена схема процессов, приводящих к фотомодификации мембранной АХЭ, учитывающая вклад УФ-превращений самого фермента, а также отдельных структурных компонентов мембран (рис. 46). Кроме того,
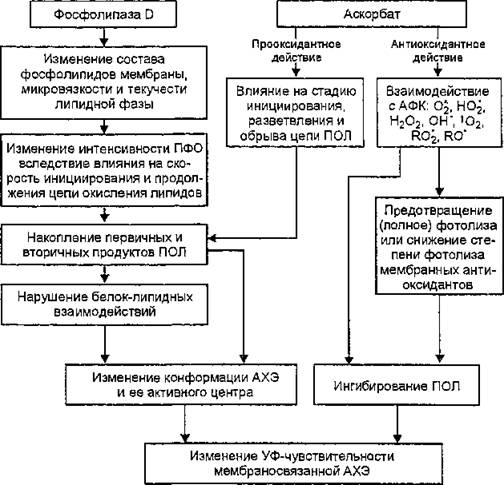
Рис. 47. Модуляция фоточувствительности мембраносвязанной ацетилхолинэстеразы
разработана схема модуляции фоточувствительности ацетилхолинэстеразы путем модификации структурного состояния липидной фазы эритроцитарной мембраны в присутствии фосфолипазы D и аскорбиновой кислоты (рис. 47).
Выявлены важная роль микроокружения в процессах функционирования АХЭ и возможность целенаправленного регулирования степени ее УФ-чувствительности путем введения в изучаемую систему экзогенных агентов, модифицирующих конформацию ближайших “соседей” фермента в биомембране. Исследования подобного рода полезны при обсуждении вопросов, касающихся выяснения механизмов влияния УФ-излучепия на структурно-функциональное состояние эритроцитарных мембран в норме и в присутствии ряда химических соединений. Они расширяют современные представления об особенностях фотохимических превращений белков-ферментов в составе мембран клеток и могут быть использованы для разработки новых физико-химических подходов к мембранному скринингу разнообразных по своей природе и механизмам действия химических соединений, а также к выявлению структурных нарушений компонентов мембран, обусловленных действием указанных внешних факторов.
4.2.3.