1.2.1. Классификация, структура и функции мембранных белков
Белки обеспечивают выполнение мембранами их специфических функций. Поэтому содержание и типы белков в различных мембранах значительно варьируют. Так, в миелиновой мембране, выполняющей функцию изолятора, белки составляют около 20 % массы мембраны.
Цитоплазматическая мембрана животных клеток на 50 % состоит из белка, во внутренней мембране митохондрий на его долю приходится около 75 % (табл. 3). Плотность мембран прямо пропорциональна содержанию в них белка.Обычно мембранные белки подразделяют на наружные (периферические) и внутренние (интегральные). При этом критерием служит степень жесткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, с низким или высоким значением pH, в присутствии растворов хелатирующих агентов (ЭДТА), связывающих двухвалентные катионы. Такие белки контактируют с поверхностью мембраны в основном за счет слабых электростатических взаимодействий с полярными головками липидных молекул либо с молекулами других белков. В определенных условиях с биомембранами могут взаимодействовать некоторые водорастворимые цитоплазматические белки. Поэтому возникла необходимость в дополнительной классификации периферических белков, которые делятся на две группы: собственно периферические и поверхностные. Собственно периферические белки, как правило, связаны с мембраной электростатическими силами и могут быть частично погружены в ее гидрофобную область. Поверхностные белки прикрепляются к мембране только за счет электростатических взаимодействий. К периферическим белкам относят гексокиназу, связанную с белком порином, локализованным во внешней мембране митохондрий; цитохром С, который образует комплекс с мембранным белком цитохромоксидазой; креатинфосфокиназу, взаимодействующую с кардиолипином внутренней мембраны митохондрий.
В группу периферических включают и белки цитоскелета: актин и миозин, ассоциированные с внутренней поверхностью мембраны в клетках эукариот, а также спектрин эритроцитов. У эукариот к числу периферических белков относят коллаген, фибро- нектин, ламинин, расположенные на внешней поверхности мембраны, входящие в состав экстраклеточного матрикса. Это гликопротеины, выполняющие сигнальную и структурные функции.По функциям периферические белки делятся на следующие группы:
1) регуляторно-сигнальные (белки экстраклеточного матрикса);
2) структурно-каркасные (актин-спектриновые комплексы);
3) белки, обеспечивающие подвижность клеток и субклеточных структур (белки микротрубочек и микрофиламентов).
Таблица З
Белковый и липидный состав некоторых мембран животных и бактериальных клеток
| Мембраны | Основные белки | Основные липиды | Отношение липид/белок по массе сухого вещества) |
| Миелин человек | Основный белок липофилин протеолипид | ФХ— 10 %, ФЭ —20 %, ФС — 8,5 %, СМ — 8,5 %, ганглиозиды —■ 26 % , холестерин — 27 % | 3—4 |
| Мембраны дисков бык | Родопсин | ФХ—41%, ФЭ — 39 %, ФС — 13 %, следы холестерина | 1 |
| Эритроциты человек | Белок полосы 3, гликофорин, спектрин, глицеральде- гид-3-фосфат- дегидрогеназа | ФХ—25 %, ФЭ — 22 %, ФС —10 %, СМ —17 %, холестерин — 25 % | 0,75 |
| Мембраны холинергического рецептора Torpedo marmorata | Ацетилхолиновый рецептор | ФХ—24 %, ФЭ — 23 %, ФС —9,6 %, холестерин — 40 % | 0,5—0,7 |
| Саркоплазма тический ретикулум кролик | Са2+-АТФаза | ФХ — 66 %, ФЭ —12,6%, ФИ — 8,1 %, холестерин — 10 % | 0,66—0,70 |
| Пурпурная мембрана Halobacterium lialobium | Бактериородоп- син | Фосфатидилглицерол- фосфат — 52 %, гликолипиды — 30 %, нейтральные липиды — 6 % | 0,2 |
Для высвобождения из мембраны интегральных белков необходимо использовать детергенты или органические растворители.
Детергенты разрушают липидный бислой и связываются с неполярными участками мембранных белков, контактирующими с гидрофобной областью бислоя. Интегральные белки представляют собой глобулярные амфифильные макромолекулы, взаимодействующие и с гидрофобными, и с гидрофильными КОМПО-нентами мембраны. Особенностью их структуры является высокое содержание а-спиральных участков и участков с конформацией статистического клубка. Степень погружения интегральных белков в липидный матрикс определяется их аминокислотным составом (количеством аминокислотных остатков с неполярными боковыми радикалами) и трехмерной пространственной структурой. Эти белки выполняют в мембране транспортную, рецепторную и ферментативную функции. К ним относят гликофорины, АТФазы, цитохром Ь5, родопсин, бактериородоп- син и др. Следует отметить, что с высоким разрешением установлена структура только бактериородопсина, однако и в этом случае положение белка относительно липидного бислоя не оп ределено однозначно. Детальное исследование интегрального мембранного белка должно включать экспериментальное изучение его топографии. Для этого используют целый арсенал методов: протеолиз, иммунологические методы (применение моноклональных и поликлональных антител против пептидов, соответствующих определенным областям белка), химическую модификацию (например, солями диазония), генетические подходы.
Различия периферических и интегральных белков определяют степень связывания их с мембраной, но не способ их прикрепления к бислою. На рис. 6 показаны способы прикрепления белков к мембране:
1 — связывание с “якорными” белками, погруженными в би- слой. Примеры: Fj-часть Ы+-АТФазы связана с Fg-частью, погруженной в мембрану; сукцинатдегидрогеназа, некоторые белки цитоскелета;
2 — связывание с поверхностью бислоя. Эти взаимодействия имеют либо электростатическую природу (а), например, миелиновый основный белок, либо гидрофобную, но практически без погружения в бислой (б), например, пируватоксидаза, фосфолипазы;
3 — связывание с помощью гидрофобного “якоря”.
Цитохром^ имеет короткий концевой сегмент из неполярных аминокислотных остатков (а). Некоторые белки используют в качестве “якоря” ковалентно связанные с ними жирные кислоты или фосфолипиды, например, щелочная фосфатаза эукариот ft?};4 — пересечение мембраны трансмембранными белками. Гли- кофорин имеет одиночный трансмембранный сегмент (а), а лак- топермеаза и бактериородопсин — несколько (б).
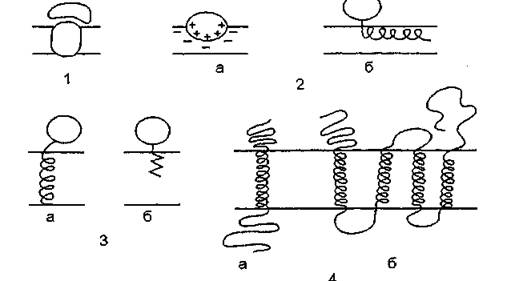
Рис. 6. Способы прикрепления белков к мембране. Объяснения в тексте
Мембранные белки эукариот могут быть ковалентно связаны с липидами. Их подразделяют на три группы:
— белки, связанные с миристиновой кислотой (14:0): каталитическая субъединица сАМР-зависимой протеинкиназы, NADPH- цитохром Ь5-редуктаза;
— белки, связанные с пальмитиновой кислотой (16:0): родопсин, анкирин;
— белки, связанные с гликозилфосфатидилинозитолом: аце- тилхолинэстераза, 5'-нуклеотидаза, щелочная фосфатаза.
Белки, связанные с жирными кислотами, по-видимому, локализованы в основном на цитоплазматической поверхности плазматической мембраны, а белки, взаимодействующие с фосфатидилинозитолом, — на наружной. Миристиновая кислота присоединяется к белку через амидную связь с N-концевым глицином, пальмитиновая — путем образования тиоэфирной связи с цистеином или гидроксиэфирной связи с серином и треонином. Эти аминокислотные остатки расположены внутри основной части полипептида вблизи трансмембранных участков, как правило, на цитоплазматической стороне. Связь белков с производными фосфатидилинозитола локализована на С-концевом участке аминокислоты. Липиды присоединяются к белку либо посттрансляционно, либо одновременно с трансляцией на рибосомах (котрансляционно).
Поверхностные белки клеток млекопитающих, а также большинство рецепторов и транспортных белков почти всегда гликози-
лированы. Олигосахаридные остатки могут защищать белки от протеолиза или участвовать в узнавании и адгезии.
Выделяют две группы олигосахаридных структур мембранных гликопротеинов:— N-гликозидные олигосахариды, связанные с белками через амидную группу аспарагина;
— О-гликозидные олигосахариды, связанные через гидроксильные группы серина или треонина.
Гликофорин А мембраны эритроцитов гликозилирован путем присоединения одного N-гликозидного олигосахарида и 15 се- рин/треонин-связанных олигосахаридов.
Наиболее детально исследован белковый состав эритроцитов млекопитающих (табл. 4).
Классической работой по электрофоретическому разделению белков мембран эритроцитов человека является исследование Фейрбанкса и соавт. (1971), в котором предложена номенклатура полипептидных полос, выявляемых в солюбилизированной с помощью додецилсульфата натрия (ДСН) мембране. Этой номенклатурой пользуется в настоящее время большинство ученых. Наличие сетчатой структуры, выстилающей внутреннюю поверхность мембраны эритроцита, было обнаружено непосредственно с помощью электронной микроскопии после обработки клеток неионным детергентом тритоном Х-100. При определенных экспериментальных условиях (в среде с высокой ионной силой и при низкой температуре) применение этого детергента позволяет полностью солюбилизировать липидный бислой и интегральные белки. При этом остается сеть белков, сохраняющая исходную форму клетки, которая представляет собой мембранный скелет (каркас) эритроцита. Он тестируется в виде двухмерной сети филаментов, длина которых зависит от особенностей приготовления препарата для микроскопии. Необходимо отметить, что белки, близкородственные компонентам цитоскелета эритроцитов, обнаружены в ряде неэритроидных клеток. В связи с универсальностью данной системы следует более подробно рассмотреть структуру и свойства некоторых цитоскелетных белков.
1.2.2.