Плазматическая мембрана
Все клетки снабжены тонкой хрупкой липопротеидной мембраной, регулирующей обмен между цитоплазмой и окружающей средой. Толщина мембраны обычно составляет 5—10 нм. Исследование тонких срезов с помощью электронного микроскопа после фиксации перманганатом показало, что мембрана состоит из трех слоев: двух плотных линий толщиной 2—3 нм с промежутком между ними в 2,5—4 нм.
Похожая структура окружает и цитоплазматические органеллы, с той лишь разницей, что мембрана ядра, митохондрий и хлоропластов двойная.Согласно современной концепции все биологические мембраны, в том числе и плазматические мембраны млекопитающих, состоят из фосфолипидного бислоя с параллельно расположенными углеводородными цепями, примерно в 16 атомов углерода длиной. Этот бислой обладает некоторыми свойствами двумерной жидкости, а именно: каждая молекула липида может перемещаться в пределах своего монослоя, но не может переходить в другой монослой. Такая липидная матрица является основной структурой мембраны. В то время как некоторые молекулы белков покрывают часть мембраны, особенно ее наружную сторону, другие белковые цепи пронизывают липидный слой насквозь и, объединившись вместе, образуют заполненные водой поры или каналы [Wallach, Zahler, 1966]. Именно эти белки ответственны за выполнение большинства функций мембраны, например прием и преобразование химических сигналов, приносимых гормонами, нейромедиаторами, факторами роста и антигенами. Именно они формируют три основных типа клеточных контактов: плотные, щелевые и синаптические. Кроме того, эти белки принимают участие в транспорте ионов и молекул.
В жидкомозаичной мембране белки свободно передвигаются в липидном слое, образуя на поверхности клетки различные структуры [Singer, Nicolson, 1972].
Некоторые белки располагаются в бислое в виде спиралевидных палочек, другие выглядят как глобули (рис.
5.5). Большинство, а возможно, и все пронизывающие мембрану белки содержат в себе молекулу углевода, расположенную на цитоплазматической стороне мембраны и выполняющую роль гидрофильного якоря для белка.В мембранах двух соседних клеток одной ткани имеется органелла, регулирующая обмен ионами и молекулами между клетками, — коннексон. Коннексон представляет собой цилиндр, образованный шестью одинаковыми белковыми субъединицами, которые могут, двигаясь, закрывать или открывать внутренний канал [Unwin, Zampighi, 1980].
К особенностям строения плазматических мембран можно отнести влияние избытка ионов кальция, обеспечивающих стабилизацию мембран и регулирующих функционирование ионных
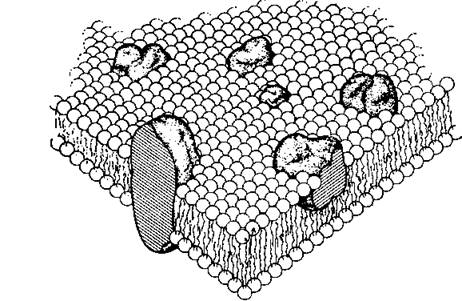
Рис. 5.5. Плазматическая мембрана.
}
каналов. Стабильность биологических мембран достаточно высока: например, при обработке фосфолипазой мембран эритро-1 цитов, митохондрий и ЭР они теряют около 70% фосфатидилхо- лина (4.59). При этом уменьшается площадь мембран, но сохраняется целостность мембранных белков и не происходит никаких конформационных изменений [Trump et al., 1970].
Липиды, в которых преобладают фосфолипиды, состоят из лецитина (фосфатидилхолин), триглицеридов (обычные жиры), жирных кислот и холестерина (химия липидов см. разд. 4.4). Фосфолипидный бислой имеет характеристичную «точку плавления», т. е. температуру, при которой в нем происходит фазовый переход из твердого состояния в жидкое. Температура этого перехода зависит от природы гидрофильных головок липидов, а также от длины и степени ненасыщенности углеводородных цепей. Липиды с длинными ненасыщенными углеводородными цепями имеют обычно и самые высокие точки плавления.
В большинстве биологических мембран липиды распределены асимметрично. Наружную поверхность бислоя составляют главным образом нейтральные липиды, а отрицательно заряженные компоненты, преимущественно фосфатидилсерин, сосредоточены на внутренней стороне.
Разность потенциалов между внутренней частью такой мембраны и омывающей ее жидкостью, измеренная с помощью калиевой нонактиновой пробы, может составлять до 300 мВ [Latorre, Hall, 1976] (о нонактине' см. разд. 14.2), что обусловливает такие типичные свойства1 мембран, как, например, воротный потенциал нервных клеток.Содержащие холестерин мембраны отличаются повышенными жесткостью и температурой плавления, так как плоские сте-1 роидные циклы интеркалируют между длинными цепями моле-1
кул фосфолипидов и ограничивают их подвижность [Coleman, 1973]. Этот вывод подтверждается и данными спектров ПМР и ЭПР (с использованием спиновых меток) искусственных фосфолипидных мембран [Gent, Prestegard, 1974].
Мембраны содержат немало различных белков, среди которых могут быть ферменты (например, аденозинтрифосфатаза) и рецепторные белки (разд. 2.1). Логично предположить, что1 белки располагаются в мембране таким образом, что большинство полярных остатков аминокислот находится вне мембраны,, а большая часть остатков неполярных аминокислот — внутри.
В различных типах мембран молярное соотношение составляющих их липидов и белков варьируется очень сильно: от одного предельного соотношения 9:1, характерного для миелиновых мембран, покрывающих нервные волокна, до другого пре- дела, когда это соотношение (в мембранах митохондрий) равно 1 : 1. В состав липидов миелиновых мембран входят холестерин, фосфатидилэтаноламин и цереброзиды (представляющие собой' не содержащие фосфора продукты конденсации этаноламина, жирных кислот и гексозы), тогда как липиды мембран митохондрий преимущественно состоят из фосфатидилэтаноламина, лецитина и кардиолипина (фосфатидилглицерин).
А. Мембраны животных клеток. Плазматические мембраны животных клеток, как правило, обогащены фосфолипидами, а именно: фосфатидил-холином, -серинам и -этаноламином'. Предполагают, что каждому мембранному ферменту для выполнения своих функций необходим определенный фосфолипид [Coleman, 1973].
В жирах этих мембран в положении 2 глицерина чаще всего находится остаток арахидоновой кислоты. В их состав входит также и холестерин. Высоковозбудимые' мембраны аксонов см. разд. 7.5.1.Б. Мембраны растительных клеток. Плазматическая мембрана растительных клеток называется плазмалеммой. В настоящее время точно установлено, что плазмалемма служит осмотическим барьером в клетке. Уникальной особенностью растительных клеток является наличие в них крупных вакуолей, большей частью заполненных водным раствором, изотоничным цитоплазме. Обычно вакуоли служат для хранения отходов. Однако в некоторых фазах роста клеток вакуоли наполняются ферментами. Тонопласт, окружающий вакуоли, по составу и свойствам, по-видимому, очень похож на плазматическую мембрану.
Во всех мембранах гриба Neurospora crassa имеются мио- инозитолсодержащие липиды. У мутантов, не содержащих инозитол, наблюдается вырождение мембран. Это свидетельствует о том, что миоинозитол необходим для сбалансированного роста этих дрожжей. Поэтому его аналоги могут останавливать их рост [Shatkin, Tatum, 1961]. Миоинозитол, вероятнее всего, находится в мембранах в виде миоинозитол-1-фосфата (4.60).
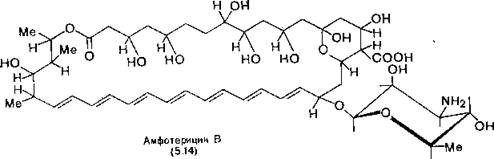
Необходимым компонентом плазматической мембраны грибов, в том числе и дрожжей, является эргостерин. Поэтому используемые в клинике полиеновые антибиотики, такие как ам- фотерицин В (5.14), повышают текучесть этих мембран и даже .разрушают их, внедряясь в мембрану вдоль стероида и нарушая тем самым ее однородность (см. также разд. 14.3).
В. Мембраны бактерий. Цитоплазматическая мембрана бактерий обладает целым рядом необычных свойств. При полном ■гидролитическом расщеплении клеточной стенки лизоцимом мембрана становится наружным слоем клетки. Ее толщина около 6—10 нм и иногда она может образовывать одиночные шпячивания в цитоплазму [Hughes, 1962]. Плазматическая мембрана бактерий составляет примерно 10% сухого вещества клетки и содержит около 25% липидов.
В других компонентах бактериальной клетки липидов почти нет. В состав М. lysode- ikticus входит около 89% фосфолипидов, это преимущественно дифосфатидилглицерин и фосфатидилинозитол. Дифосфатидил- іглицерин содержит главным образом разветвленные алифатические кислоты с цепью из 15 углеродных атомов [Macfarlane, ■1961]. Стероидов бактерии не содержат.В мембранных белках найдены только обычные аминокислоты. Из плазматических мембран различных бактерий хроматографически на липофильном сорбенте было выделено много белковых фракций, и оказалось, что к одной из них присоединена ДНК одиночной бактериальной хромосомы [Daniels, 1971]. Юбычным компонентом мембран является РНК [Hughes, 1962].
Плазматические мембраны бактерий содержат также пер- ;меазы и ферменты, синтезирующие клеточную стенку [Crathorn, Hunter, 1958]. Из-за небольших размеров бактерий в них нет митохондрий, поэтому большинство митохондриальных ферментов расположено на плазматической мембране. Такое открытое расположение многих жизненно важных ферментов повышает уязвимость бактерий к действию избирательно токсических веществ (см. также разд. 5.4.3).
В отличие от бактерий у другого класса прокариот — мико- плазм, не имеющих клеточных стенок, стероиды (чаще всего эргостерин) — это основные компоненты, обеспечивающие целостность мембраны. 'У растений и животных микоплазмы вызывают различные заболевания, легко поддающиеся лечению полиеновыми антибиотиками и тетрациклинами.
Г. Мембраны раковых клеток. Превращение здоровой клетки в раковую сопровождается отчетливыми изменениями структуры ее поверхности. Как было показано в экспериментах с внутриклеточным введением электродов, клетки образуют упорядоченную ткань благодаря своей способности чувствовать присутствие соседних клеток и с ними обмениваться информацией через плазматические мембраны [Loewenstein, Kanno, 1967]. Однако в злокачественных клетках отсутствует механизм регуляции роста клеток (см. также разд. 5.1)..
Повышенное содержание сиаловой кислоты (5.15) на поверхности злокачественных клеток (например, лейкозных и лимфосаркомы человека) приводит не только к изменению их гликопротеинового состава, но и к видимым, структурным изменениям [Van Веек, Smets, Emmelot, 1975].
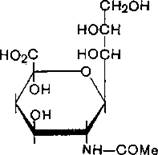
N-Ацетилнейраминовая кислота- (сиаловая нислота)
(5.15)
Д. Искусственные мембраны. На этих мембранах выполнено много исследований по изучению возбудимости мембран [Mueller, Rudin, 1967] и их проницаемости для ионов. Наиболее' удобным методом получения искусственных мембран является обработка ультразвуком смеси лецитина с водой. При этом образуется суспензия везикул с бислойной структурой [Huang, 1969]. На таких везикулах можно изучать транспорт веществ- через природные мембраны.
Взаимодействие и движение белков и липидов в мембранах, динамика мембран см. Houslay, Stanley (1982); структура m функция мембран см. Gomperts (1976), Chapman (1968—1982, 4 тома), а также Finean, Coleman, Michell (1978)'.
5.4.2.