Нарушение структурно-функциональных свойств клеточных мембран
Одним из наиболее важных структурных компонентов клеточной мембраны являются фосфолипиды, так как именно им принадлежит главная роль в формировании параметров проницаемости бислоя, в регуляции активного и пассивного транспорта органических веществ и электролитов, обеспечении чувствительности к действию гормонов и регуляции активности ферментативных систем [198].
Все фосфолипиды имеют общую черту — наличие в молекуле как полярных групп («головок»), так и неполярных гидрофобных углеводородных цепей жирных кислот (от 12 до 24 атомов углерода), что обеспечивает им амфифильность, а наличие цис-конформации двойной связи в ненасыщенных жирных кислотах придает слою жидкостность, менее упорядоченную структуру, чем в случае насыщенных или транс-ненасыщенных жирных кислот (рис. 22).Достаточно большое внимание в современных исследованиях уделяется процессам дестабилизации мембранных структур под влиянием активных форм кислорода [382]. Показано, что у больных сахарным диабетом типа II перекиси полиненасыщенных жирных кислот, включающиеся в составе фосфолипидов в структуру клеточных мембран эндотелия и форменных элементов крови, значительно изменяют ее, приводя к еще большему повреждению функционально неактивных гликозилированных белков, углеводов и липидов, служащих мишенью для ПОЛ [163]. Фосфолипиды, образующие бислойную липидную мембрану, вследствие своей высокой ненасыщенности являются преимущественным субстратом, подвергающимся повреждающему действию АКМ [121]. Основой всех модифицирующих или повреждающих эффектов ПОЛ в мембране является реакция ненасыщенных жирнокислотных остатков фосфолипидов бислоя
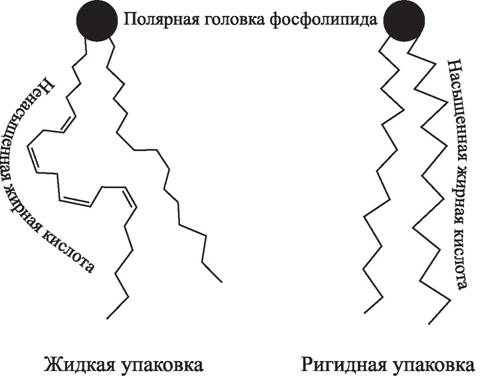
Рис. 22. Структура фосфолипидов
1 110 г
с активными формами кислорода, в результате которой в молекулах фосфолипидов, содержащих во втором положении жирную кислоту, появляется полярная ОН-группа.
Образующиеся в результате включения полярной группировки гидроперекиси фосфолипидов неустойчивы и, соответственно, их накопление приводит к цепи дальнейших превращений.Фосфолипиды не являются статичными структурными компонентами мембраны. Они находятся в динамическом равновесии внутри бислоя, и этот плеоморфизм липидов лежит в основе физико-биохимических свойств мембраны [803]. Трансмембранная фосфолипидная асимметрия — характерное свойство биологических мембран [1766]. Наружный слой мембран сформирован преимущественно из холинофосфолипидов — фосфатидилхолина и сфингомиелина, тогда как аминофосфолипиды — фосфатидилсе- рин и фосфатидилэтаноламин — удалены к цитоплазматической поверхности бислоя. Подобная асимметрия поддерживается кооперацией трех транспортеров:
• АТФ-зависимой аминофосфолипид-специфической транслоказой (флиппа- зой), которая способна быстро перемещать фосфатидилсерин и фосфатидилэтаноламин от наружного к внутреннему слоям мембраны против градиента концентрации;
• АТФ-зависимой неспецифической липидфлоппазой, которая медленно транспортирует липиды от внутреннего к наружному слоям мембраны;
• Са2+-зависимой неспецифической липидскрамблазой, которая позволяет липидам случайно перемещаться между обоими слоями мембраны [1850].
При физиологических концентрациях Са2+ асимметричное распределение фосфа- тидилсерина поддерживается активностью транслоказы и флоппазы в условиях инактивации скрамблазы. Повышение уровня цитоплазматического Са2+ непосредственно активирует последнюю, вызывая хаотичное перемещение фосфатидилсерина внутри мембраны [1766]. Появление фосфатидилсерина на наружной поверхности мембраны способствует коагуляции и тромбозу, так как предоставляет каталитическую поверхность для образования протромбиназного теназного комплекса. Кроме того, такие клетки распознаются фагоцитами как патологическая цель для элиминации, что является физиологическим механизмом удаления состарившихся эритроцитов из системы кровообращения, поскольку эритроциты имеют свойство аккумулировать небольшие количества поверхностно-расположенного фосфатидилсерина на протяжении жизненного цикла [1850].
Таким образом, при физиологических концентрациях Са2+ АТФ-зависимые транслоказа и липидфлоппаза взаимодействуют для поддержания устойчивого динамического асимметричного равновесия, при котором все фосфолипиды медленно и непрерывно движутся к наружной поверхности мембраны, тогда как аминофосфолипиды переправляются назад — к внутренней поверхности бислоя. При высоких же концентрациях цитоплазматического Са2+ происходит блокада этих ферментов, активация неспецифической липидскрамблазы, что приводит к хаотическому распределению фосфолипидов в бислое [1766]. Очевидно, что промежуточные концентрации Са2+ приводит к ситуации, когда функционируют оба механизма, составляя оппозицию друг другу. При этом происходит устойчивое перераспределение мембранных фосфолипидов, характеризующих конкретную патологию.
По всей видимости, основным механизмом нарушения фосфолипидной асимметрии при сахарном диабете является перекисное окисление мембранных липидных транспортеров, приводящее к снижению как АТФ-зависимой ферментативной активности, так и Са2+-насоса [377]. В создавшихся условиях происходит повышение концентрации цитоплазматического Са2+, стимулирующее скрамблазу и еще более подавляющее транслоказу. Таким образом, нарушение окислительных процессов в мембране приводит к экспозиции фосфатидилсерина на ее поверхности, снижению продолжительности жизни эритроцита, снижению деформируемости мембраны, повышению ее вязкости, повышению спонтанной агрегации и адгезии к эндотелиальным клеткам [1711, 1766]. Последние исследования показали, что фосфатидилсеринзависимое распознавание клеток не ограничивается компетенцией фагоцитов и что гладкомышечные клетки также способны узнавать клетки, имеющие на своей поверхности фосфатидил- серин. При этом происходит их адгезия к субэндотелиальным структурам с формированием первичной гемостатической бляшки [1850]. Присутствие фосфатидилсерина на поверхности агрегировавших тромбов контролирует формирование тромбина, поскольку мембраны таких тромбоцитов предоставляют каталитическую поверхность как для прокоагулянтных, так и для антикоагулянтных реакций [1766].
Все вышеперечисленное вновь и вновь подтверждает роль свободнорадикального окисления в дестабилизации клеточных мембран как одного из основных механизмов развития диабетической ангиопатии.
Существует большое количество данных, подтверждающих прямое и косвенное участие гемореологических факторов в развитии микрососудистых и атеросклеротических осложнений при сахарном диабете типа II [811, 1711]. При исследовании эритроцитов выявлен целый ряд характерных нарушений: снижена продолжительность жизни эритроцитов, обнаружено изменение фосфолипидной асимметрии, повышена агрегация и вязкость, снижена способность мембраны к деформации, увеличена адгезия к эндотелиальным клеткам [814, 1380].
Wali et al. показали, что у больных диабетом эритроциты в 2—3 раза сильнее прилипают к культуре человеческих эндотелиальных клеток, чем эритроциты здоровых людей, и что это повышение адгезивной способности не связано с гипергликозилиро- ванием поверхностных белков, поскольку нормальные эритроциты, подвергнутые инкубации с концентрированным раствором глюкозы (200 мг/дл) в течение 20 часов, не демонстрировали повышение адгезии [1711, 1766]. Авторы связывают это нарушение с изменением организации фосфолипидов в бислое эритроцитарной мембраны, что проявляется повышением содержания фосфатидилсерина и фосфатидилэтаноламина в наружном слое и менее упорядоченной, т. е. более рыхлой упаковкой липидов на поверхности мембраны. Однако отмеченные эффекты обусловлены не статичным повышением содержания фосфатидилсерина и фосфатидилэтаноламина в наружном слое мембраны, а повышением скорости транслокации этих фосфолипидов между слоями. Помимо этого, экстернализация фосфатидилсерина может объясняться окислением белков цитоскелета [463].
Интересно, что изменение структурно-функциональных свойств клеточной мембраны может не только лежать в основе гемореологических расстройств, но и приводить к возникновению инсулинрезистентности [407, 962]. Это связано с тем, что фосфолипидный состав определяет жидкостность мембраны, от которой зависит инсулин-ре- цепторное связывание и передача сигнала.
В нескольких исследованиях in vivo выявлена положительная корреляция между жидкостностью мембраны и связыванием и (или) действием инсулина [407]. В ряде работ была продемонстрирована четкая связь между жирнокислотным составом мембранных фосфолипидов скелетных мышц и действием инсулина [546, 1800]. Показано, что при инкубации скелетных мышц с пальмитиновой, олеиновой, линолевой и линоленовой кислотами снижается инсулин-стимулиро- ванный синтез гликогена на 51 % по сравнению с контролем. Кроме того, инкубация с олеиновой и линолевой кислотами снижает инсулин-стимулированный транспорт и фосфорилирование глюкозы [1623]. Другие исследователи отмечали увеличение количества фосфатидилэтаноламина и общих фосфолипидов в мембранах эритроцитов у больных с нарушенной толерантностью к глюкозе по сравнению с пациентами с нормальной толерантностью к углеводам [407]. Уровень инсулинрезистентности достоверно коррелировал с индексом массы тела и содержанием сфингомиелина в мембранах эритроцитов. Кроме того, в этом исследовании была показана зависимость между повышением уровня инсулина плазмы натощак и увеличением содержания сфингомие- лина в мембране эритроцита.Выявленные у больных сахарным диабетом дестабилизация липидного бислоя и ингибирование большинства мембраносвязанных ферментов во многом обусловлены активацией в клеточных мембранах окислительных процессов; это делает необходимым более глубокое изучение мембранологических аспектов диабетологии и поиска способов коррекции выявленных нарушений [733, 1695].