Цитоскелет (мембранный каркас)
Основными компонентами мембранного скелета эритроцита являются спектрин, актин, анкирин, а также белки полосы 4.1 и 4.9. Мембранный скелет содержит около 50 % всех белков, присутствующих в мембране.
Таблица 4
Свойства, степень ассоциации и функции некоторых эритроцитарных мембранных белков
| Пептидная фракция | Молекулярная масса субъединицы | Число молекул на клетку, х 10~3 | Степень ассоциации | Функция | Связь с белками |
| Полоса 1 (спектрин) Полоса 2 (спектрин) | 240000— 260000 (а) 220000 (Р) | 2,2 | Димеры полос 1 и 2 образуют тетрамеры и олигомеры | Участвует в формировании мембранного скелета | Полоса 3 (с полосой 1), анкирин (с полосой 2), полоса 4.1 |
| Полоса 2.1 (анкирин) | 215000— 166000 | Мономер | Связывает спектрин с мембраной через белок полосы 3 | Спектрин, полоса 3 | |
| Полоса 3 | 89000—95000 | 12 | Тетрамер в равновесии с димерами | Транспорт неорганических анионов, взаимосвязь с мембранным скелетом | Анкирин, полосы 4.1 и 4.2, гликоли- тические ферменты |
| Полоса 4.1 | 78000—80000 | 2,3 | Димер | Связывает мембранный, скелет с инте гральными белками; стабилизирует спект- рин-актиновые взаимодействия | Спектрин, гликофорин |
| Полоса 4.5 | 52000—59000 | 0,7—1,4 | ? | Системы транспорта моносахаридов и нуклеозидов | ? |
| Полоса 4.9 | 45000—48000 | 1 | ? | Связывание актина | Актин |
| Полоса 5 (актин) | 43000 | 5,1 | Олигомеры из 12—17 субъединиц | Участвует в формировании мембранного скелета | Спектрин, полоса 4.1 |
| Полоса 6 | 35000 | 4,1 | Тетрамер | Глицераяьде- гид-3-фосфатде- гидрогеяаза | Полоса 3 |
| Гликофо- рин А Гликофо- рин В Гликофо- рин С | 29000—31000 23000 25000—29000 | 2 | Димер | Место прикрепления мембранного скелета | Полоса 4.1 |
Спектрин (рис.7), по данным электронной микроскопии» является гибкой молекулой в виде тяжа длиной 100 нм и представляет собой гетеродимер, составленный из двух полипептид- ных цепей аи|3, уложенных антипараллельно. Однако в эритроцитах in situ основной формой спектрина является тетрамер, образованный путем самоассоциации гетеродимеров конец в конец.
На обоих хвостовых концах тетрамера имеются центры связывания актина и белка полосы 4.1. Путем латеральной ассоциации тетрамер спектрина прикреплен к актину, присутствующему в интактной клетке в виде олигомеров из 12—17 субъединиц. Каждый такой протофиламент актина соединен в среднем с 6 тетрамерами спектрина, которые другим концом связаны с соседними протофиламентами актина. Предполагают, что белок полосы 4.1 облегчает ассоциацию спектрина с актином.Спектрин имеет также центры связывания анкирина, находящиеся на р-субъединице и расположенные на расстоянии 20 нм от центра тетрамера. С помощью этого якорного белка актин- спектриновая сеть прикреплена к мембране через цитоплазматический участок белка полосы 3. По-видимому, это основной механизм прикрепления каркаса к мембране. Цепь белок полосы 3 — анкирин — спектрин представляет собой путь, связывающий наружную и внутреннюю поверхности мембраны эритроцита, позволяющий осуществлять контроль за протеканием внутриклеточных событий на уровне организма (см. главу 2), Установлено, что поддержание нормальной структуры мембранного каркаса эритроцита возможно при определенном оптимальном содержании АТР и двухвалентных катионов (прежде всего Са2+) внутри клеток. По-видимому, степень фосфорилирования ком-
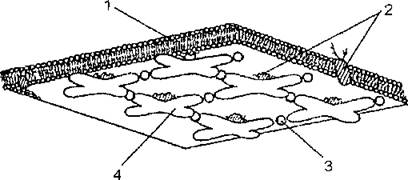
Рис. 7. Схема организации спектрин-актиновой сети: 1 — липидный бислой; 2 — интегральные белки; 3 — актин; 4 — тетрамер спектрина
3. Заказ 3788
33
понентов мембранного скелета определяет сродство Са2+ к мембранным белкам. В свою очередь, ионы Са2г обусловливают прочность связи отдельных компонентов мембранного каркаса. В связи с этим регулирование свойств последнего, а, значит, и целой клетки, может осуществляться за счет изменения соотношения [АТР]/[Са2+].
Мембранный скелет эритроцита — наиболее полно охарактеризованная разновидность цитоскелета эукариотической клетки.
В прокариотических клетках наличие цитоскелета не выявлено. В целом цитоскелет эукариот представляет собой сложную сеть волокон, обеспечивающих механическую опору для плазматической мембраны, определяющих форму клетки, местоположение клеточных органелл и их перемещение при делении клетки. Цитоскелет образуют три типа волокон:1) микрофиламенты (диаметр около 6 нм), состоящие из актина и связанных с ним белков;
2) промежуточные филаменты (диаметр 8—10 нм), состоящие из кератинов;
3) микротрубочки (диаметр 23 нм), состоящие из тубулина.
По-видимому, микрофиламенты участвуют в таких мембранных процессах, как рецепторзависимый эндоцитоз, пэтчинг и кэп- пинг антигенов на клеточной поверхности, цитокинез, клеточная подвижность. Микротрубочки образуют цитоплазматическую сеть, связывающую плазматическую мембрану с различными субклеточными компонентами.
Следует отметить,что одной из задач мембранологии является выявление молекулярных механизмов взаимодействия отдельных компонентов цитоскелета клетки и путей его регулирования, которые обеспечивают нормальное функционирование биосистем.
1.2.3.