5.1. КРАТКИЙ ОБЗОР ГРУПП МЕТОДОВ ИССЛЕДОВАНИЯ БИОМЕМБРАН
Методы исследования биологических мембран весьма разнообразны и условно объединены в 5 групп: биохимические, физиологические, иммунологические, генетические, биофизические.
Биохимические методы позволяют разделять, выделять и анализировать в чистом виде липидные и белковые компоненты, изучать их физико-химические свойства в свободном состоянии и в составе надмолекулярных комплексов в условиях воздействия различных внешних факторов (температуры, концентрации водородных ионов и др.), исследовать их время «жизни», пути биосинтеза и распада этих компонентов.
К ним относят методы выделения (недеструктивные и включающие разрушение клеток); разделения субклеточных фрагментов (хроматография, электрофорез, центрифугирование, иммуноаффинные методы); идентификации и оценки чистоты субклеточных фракций; выделения органелл и мембранных систем; экстракции липидов и разделения их по классам; количественного определения фосфолипидов; исследования трансмембранного распределения липидов; солюбилизации мембранных белков, их реконструкции и определения функциональной активности реконструированных мембран, выделения и модификации мембранных белков.Физиологические методы используют для изучения функционирования естественных и искусственных мембран. Они позволяют исследовать проницаемость мембран, процессы возбуждения, торможения, проведения нервного импульса, распределения и выведения иоЬов и молекул из клеток и тканей, изменения физиологических функций клеток.
Иммунологические методы широко используются для идентификации мембранных компонентов, их локализации, оценки количества, выделения. Это методы получения поликлональных и моноклональных антител к компонентам мембран, иммуно- блоттинг (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия с перенесением разделенных белков на нитроцеллюлозные фильтры для последующего выявления с помощью антител), иммунологическая очистка субклеточных фракций, иммунологическое выделение мембранных компонентов, использование антител для отбора комплементарных ДНК, кодирующих мембранные белки.
Генетические методи основаны на использовании мутантов, дефектных по синтезу определенных мембранных белков. Они позволяют исследовать функции мембранных белков, их роль в функционировании мембран, проблемы самоорганизации мембран.
Биофизические методы позволяют изучать динамическую организацию биомембран, получить представления об упаковке и движении липидных молекул в природных и модельных мембранах, их взаимодействии друг с другом и молекулами белков, исследовать фазовые переходы и другие процессы. К ним относятся дифракционные методы (рентгеновская дифракция, дифракция нейтронов), резонансные методы, метод электронной микроскопии, оптические методы (круговой дихроизм, дисперсия оптического вращения, абсорбционная спектроскопия, люминесценция, метод флуоресцентных зондов), метод дифференциальной сканирующей микрокалориметрии, метод моделирования и получения искусственных мембран и др.
Дифракционные методы основаны на взаимодействии электромагнитного излучения или частиц с длиной волны, соизмеримой с межатомным расстоянием, и компонентов мембраны. Их используют для определения геометрических параметров структуры: типа липидной мезофазы и ее периодичности, толщины бислоя, среднего расстояния между углеводородными цепями. К этим методам относятся рентгеновская дифракция (рентгеноструктурный анализ) и дифракция нейтронов.
В основе рентгеноструктурного анализа лежит взаимодействие рентгеновского излучения с электронами вещества, в результате которого возникает дифракция рентгеновских лучей с длиной волны - 0,1 нм. Последние рассеиваются на электронных оболочках атомов. Интерференция волн, рассеянных веществом, приводит к возникновению дифракционной картины, что позволяет зарегистрировать рентгенограмму. При рассеянии на кристалле можно рассматривать дифракцию как отражение рентгеновских лучей плоскостями кристаллической решетки. Дифракция наблюдается, если рассеянные волны находятся в фазе, т.е. разность хода лучей равна целому числу волн п.
Условие дифракции (отражения) описывает формула Брэгга—Вульфа:
пХ = 2 d sin 0,
где X — длина волны; d — расстояние между кристаллическими плоскостями; © — угол между направлением падающего луча и кристаллической плоскостью.
На основании дифракционной картины, получаемой для рентгеновских лучей с известной длиной волны, определяют параметр d. Дифракционная картина зависит от длины волны рентгеновских лучей и строения объекта. Анализ дифракционных максимумов позволяет установить распределение электронной плотности в кристалле. Рентгеноструктурный анализ дает информацию о расположении атомов в молекулах и кристаллах.
Для построения профилей электронной плотности, а также для изучения ориентации и характера упаковки углеводородных цепей используют мультислои липидов или смесей липидов и белков, ориентированные на твердой подложке. Их получают путем постепенного нанесения липидов при многократном прохождении стеклянной пластины сквозь монослой липида на границе раздела вода — воздух или путем спонтанного образования мультислоев, параллельных подложке, при испарении органического растворителя из капли липидного раствора.
Метод рентгеновской дифракции позволяет установить наличие бислойной структуры в природных и модельных мембранах и определить параметры бислоя. На основании анализа дифракционной картины можно исследовать состояние углеводородных цепей мембранных липидов и зарегистрировать фазовый переход из гелеобразного в жидкокристаллическое состояние. Однако из-за высокой лабильности и сравнительно малой упорядоченности структуры биомембран метод рентгеноструктурного анализа не позволяет изучить локализацию мембранных белков. Лишь в мембранах, содержащих упорядоченные белковые комплексы, удается выявить наличие и ориентацию а-спиральных участков интегральных белков.
С помощью метода дифракции нейтронов, дополняющего рентгеноструктурный анализ, определяют расположение молекул и их частей в бислое путем изотопного замещения водорода на дейтерий.
В отличие от рентгеновского излучения, нейтроны практически одинаково эффективно рассеиваются легки-ми и тяжелыми атомами. Взаимодействие с некоторыми ядрами, имеющими соответствующие резонансные уровни энергии, приводит к изменению знака амплитуды рассеяния. Амплитуда когерентного рассеяния ядром (Ь) связана с эффективным сечением (о) соотношением: b = д/сМя . Различие в рассеянии нейтронов атомами водорода и дейтерия позволяет широко использовать эффекты изотопного замещения для изучения биомембран.
Резонансные методы анализа веществ подразделяют на метод ядерного магнитного резонанса (ЯМР) и метод электронного парамагнитного резонанса (ЭПР).
Метод ЯМР основан на резонансном поглощении в сильном внешнем магнитном поле энергии электромагнитного радиочастотного поля системой атомных ядер, обладающих магнитным моментом. Он позволяет получать сведения о подвижности молекул липидов биомембран, об упаковке фосфолипидных молекул в бислое, используется для регистрации изменений значения pH в частицах малого размера (искусственные мембраны, митохондрии), изучать процессы латеральной диффузии липидов и трансбислойного движения (“флип-флоп”-переходы) молекул.
Положение спектральной линии ЯМР относительно некоторой эталонной линии называют химическим сдвигом. Химический сдвиг (5) — это отношение сдвига частоты поля (Доэ) к эталонному значению (о>0), умноженное на 10е:
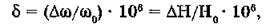
где Но — напряженность постоянного магнитного поля.
Величина 8 для алифатических протонов варьирует от -0,5 до -2,0, ароматических протонов — от -6,0 до -8,5. В качестве эталона для протонного резонанса органических соединений используют тетраметилсилан (CH3)4Si, а для водных растворов биополимеров — 2,2-диметил-2-силанпентан-5-сульфоновую кислоту (ДСС). При высоком разрешении наблюдается сверхтонкая (мультиплетная) структура линий ЯМР, возникающая вследствие магнитного взаимодействия между ядрами, передаваемого через электроны связи (непрямое спин-спиновое взаимодействие).
К недостаткам метода ЯМР следует отнести низкую чувствительность (концентрация образца должна быть не менее 10~а моль/л), а также малую эффективность в изучении крупных молекул, т.е. мембранных белков.
Метод ЭПР — это резонансное поглощение энергии электромагнитных колебаний в сантиметровом или миллиметровом диапазоне длин волн веществами, содержащими парамагнитные частицы (молекулы, атомы, ионы, свободные радикалы, слабо связанные с атомом электроны). Условие резонанса, при соблюдении которого образец поглощает электромагнитную энергию, описывают уравнением
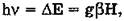
где h — постоянная Планка; v — частота переменного электромагнитного поля; АЕ — разность энергий между магнитными подуровнями, которые образуются в результате расщепления уровней энергии парамагнитной частицы в постоянном магнитном поле Н; g — фактор расщепления (отношение магнитного момента парамагнитной частицы к ее механическому моменту (спину); Р — магнетон Бора.
Метод ЭПР основан на введении в липидный бислой парамагнитных меток и зондов. Основу спиновых меток и зондов составляет стабильный свободный иминоксильный (нитроксильный) радикал с неспаренным электроном, локализованным преимущественно у атома азота:
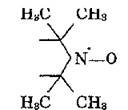
Если производное такого радикала присоединяется к белку или липиду ковалентной связью, то такое производное называют спиновой меткой, а если с помощью электростатических сил и гидрофобных взаимодействий —-то спиновым зондом.
Выбор спинового зонда или метки определяется задачами исследования. Так, для изучения характера движения липидных молекул используют липофильные спин-меченные жирные кислоты, глико- и фосфолипиды. Последние применяют также для исследования латерального фазового разделения липидов и липид-белковых взаимодействий.
Некоторые амфифильные зонды, в частности 2,2,6,6-тетра- метилпиперидин-М-оксил (TEMPO), растворяются частично в углеводородной зоне мембраны, а частично — в ее полярной области, Поэтому их спектры зависят от полярности окружения.
Распределение такого зонда в мембране определяется фазовым состоянием липидов и “чувствительно” к их фазовым переходам.Гидрофильные спиновые зонды, представляющие собой модификацию зонда TEMPO и изменяющие свой заряд в зависимости от величины pH, способны связываться с различными участками полярных групп мембраны за счет электростатических взаимодействий. Их используют для исследования поверхностного потенциала и плотности заряда мембранной поверхности.
Более предпочтительным для изучения биомембран является одновременное применение нескольких зондов, по-разному взаимодействующих с компонентами мембран, что позволяет исследовать цитоплазматическую и наружную поверхности мембран клеток и клеточных органелл.
Таким образом, метод ЭПР применяют для изучения фазовых переходов в липидном бислое, микровязкости мембран, подвижности углеводородных цепей, латеральной диффузии и “флип- флоп”-переходов. Недостаток этого метода заключается в том, что введение зонда изменяет структуру бислоя и свойства мембраны. Метод ЭПР более чувствителен по сравнению с методом ЯМР, так как магнитный момент электрона в 1000 раз выше, чем ядра.
С помощью метода электронной микроскопии (разрешающая способность электронного микроскопа находится в диапазоне 2— 4 нм и превышает таковую для оптического микроскопа более чем в 500 раз) исследуют морфологию внутренних поверхностей двух монослоев, распределение фаз в мембране, трехмерную молекулярную структуру мембранных белков, форму и размеры солюбилизированных мембранных белков. Однако необходимость проведения обезвоживания, фиксации и контрастирования объекта солями тяжелых металлов приводит к существенной модификации структурного состояния компонентов мембран.
Разновидность метода электронной микроскопии — “замораживание—скалывание”. Его применяют для изучения характера связывания белков с мембранами. Для этого образец, не требующий химической фиксации, замораживают в жидком азоте, а затем раскалывают в плоскости наименьшего сопротивления холодным ножом в вакууме. Плоскость наименьшего сопротивления проходит между двумя слоями липидной фазы мембраны. Далее поверхность образца покрывается платиной и углеродом, его растворяют и отпечаток (реплику) рассматривают в электронный микроскоп. Бели лед возгоняют с поверхности образца, то модификацию метода называют “замораживание—травление”.
Оптические методы позволяют получить информацию о механизме фотосинтеза, электронном транспорте, транспорте кислорода в тканях, транспорте ионов, взаимодействии веществ различной природы с мембранами, белок-липидных взаимодействиях и других процессах. Они основаны на присутствии в изучаемой системе эндогенных или экзогенных (вносимых в систему экспериментатором) хромофорных групп. К эндогенным хромофорам относятся порфирины, флавины, каротиноиды, пиридиннуклеотиды, цитохромы, гемоглобин, миоглобин, которые поглощают свет в видимой области спектра. Акридины, нафталин- сульфонаты, цианины являются экзогенными хромофорами. К оптическим методам относят абсорбционную спектрофотометрию, люминесценцию, метод флуоресцентных зондов, а также круговой дихроизм, дисперсию оптического вращения. Последние наряду с ИК-спектроскопией и спектроскопией комбинационного рассеяния используются для определения содержания различных элементов вторичной структуры молекулы белка, позволяют изучать ее конформационные переходы.
Люминесценция наблюдается в результате поглощения веществом энергии возбуждения, перехода его частиц из основного в возбужденное электронное состояние и обратно. Она представляет собой свечение атомов, ионов, молекул и их комплексов, возникающее в результате электронного перехода из возбужденного состояния в основное.
Все известные виды люминесценции подразделяют на два больших класса: флуоресценцию и фосфоресценцию. Под флуоресценцией понимают свечение, мгновенно (10“в с) затухающее после прекращения возбуждения. Квант флуоресценции испускается при переходе из синглетного возбужденного состояния Sj (с его нижнего колебательного подуровня) в синглетное основное состояние So: St* —» So + hv^. Фосфоресценция — свечение, продолжающееся в течение длительного промежутка времени (>10'с с) после прекращения возбуждения. Квант фосфоресценции высвечивается при переходе (с обращением спина) электрона из триплетного возбужденного состояния Т, в основное состояние: Т, -> Sn + hv^. .
1 10 фас
Есть и другие виды классификации люминесценции: по типу возбуждения свечения и кинетике процесса люминесценции. Так, в случае возбуждения вещества световыми квантами возникающее при этом свечение называется фотолюминесценцией.
В зависимости от кинетических характеристик свечения выделяют резонансную, спонтанную, вынужденную и рекомбинационную люминесценцию.
Резонансная флуоресценция — основа процесса атомной люминесценции — представляет собой излучение фотонов той же энергии, что и у поглощенных фотонов возбуждающего света; наблюдается в газах и кристаллах.
При спонтанной люминесценции после возбуждения молекула сначала переходит на высокий возбужденный электронный уровень S2, а затем путем безызлучательного перехода — на более низкий возбужденный уровень Sx. Квант люминесценции высвечивается при переходе Sj So, поэтому его величина оказывается меньше, чем у поглощенного кванта. Спонтанная люминесценция наблюдается у паров, растворов сложных молекул, молекулярных кристаллов.
Вынужденная люминесценция характерна для сложных органических молекул, находящихся при низкой температуре или в жестких средах. В этом случае при возбуждении молекула переходит из состояния So в возбужденные состояния S2* и б/, затем безызлучательным путем — в Т (триплетное), переход из которого в So запрещен правилами отбора. За счет внутренней колебательной энергии или сообщенной извне тепловой энергии осуществляется переход в основное состояние с испусканием кванта люминесценции.
Спонтанный и вынужденный типы излучения наиболее характерны для молекулярных систем и поэтому их называют молекулярной люминесценцией.
Рекомбинационная люминесценция наблюдается у газов и сложных неорганических кристаллов в случае рекомбинации радикалов или ионов с образованием их возбужденных молекул.
Спектром люминесценции вещества называют зависимость интенсивности люминесценции от длины волны (частоты) измеряемого света. Спектр возбуждения люминесценции — это зависимость интенсивности люминесценции от длины волны возбуждающего света.
Теоретически спектры возбуждения люминесценции молекул должны быть идентичны по форме их спектрам поглощения и не зависеть от длины волны измерения флуоресценции. Однако чаще всего на практике эти спектры отличаются вследствие различий физико-химических свойств молекул в основном и возбужденном состояниях. Например, в возбужденном состоянии могут изменяться геометрические параметры, межатомные расстояния и, следовательно, дипольные моменты молекул, степень диссоциации последних. Для устранения кажущихся различий между спектрами поглощения и возбуждения необходимо соблюдение следующих условий: а) одинаковой интенсивности возбуждающего света во всем диапазоне частот; б) отсутствие вторичного поглощения исследуемого излучения.
Форма спектра люминесценции (правило Каши) и квантовый выход (закон Вавилова) не зависят от длины волны возбуждающего света. Это обусловлено тем, что при поглощении больших квантов возбуждающего света происходит переход молекул из основного состояния So в возбужденные S2*, S3*, однако излучательный переход осуществляется только из состояния S/ (люминесценция), а переходы S2* —> S/, S3* -> S/ происходят безызлуча- тельно.
Согласно правилу Стокса частоты возбуждающего света всегда больше или равны частотам люминесценции: т. е.
одна часть поглощаемой молекулой энергии идет на возбуждение люминесценции, а другая расходуется на увеличение ее колебательной энергии и развитие безызлучательных переходов. Однако строгое выполнение правила Стокса наблюдается у атомов и простых молекул в газовой фазе. Спектры поглощения и люминесценции молекул могут перекрываться, т.е. значения частот переходов между электронными уровнями при поглощении могут быть меньше, чем при испускании. Следовательно, наблюдается нарушение правила Стокса. Часть спектра люминесценции, где выполняется правило Стокса, называется стоксовой областью, а где оно нарушается— антистоксовой. Поэтому это правило характеризует одиночный акт поглощения и испускания света молекулой.
В соответствии с законом Стокса—Ломмеля спектр излучения и его максимум всегда сдвинуты по сравнению со спектром поглощения и его максимумом в сторону более длинных волн:
hv люм < hv 1ШГЛ
макс макс
Этот закон носит статистический характер и описывает поглощательные и излучательные свойства совокупности молекул исследуемого образца. Поэтому образование антистоксовой области спектра люминесценции не противоречит закону сохранения энергии: колебательная энергия молекул частично преобразуется в энергию их люминесценции, что удовлетворяет условию: hv > hv
Л ЮМ ПСІГЛ
Закон Стокса—Ломмеля является качественным выражением правила зеркальной симметрии спектров поглощения и люминесценции Левшина, которое гласит: спектры поглощения и люминесценции зеркально симметричны относительно прямой, проходящей перпендикулярно к оси частот (длин волн) через точку пересечения спектров.
Эффективность процесса превращения энергии возбуждающего света в энергию люминесценции характеризуется квантовым выходом люминесценции. Это отношение числа высвеченных квантов люминесценции к числу поглощенных квантов возбуждающего света.
Молекулы люминесцирующих веществ оптически анизотропны, поэтому люминесцентное излучение каждой молекулы частично поляризовано. Если анизотропные молекулы ориентированы хаотично, то вещество в целом становится изотропным, а его люминесценция — неполяризованной. При возбуждении вещества линейно-поляризованным светом его поглощение осуществляется молекулами, у которых поглощающий осциллятор параллелен электрическому вектору падающего света. Поэтому поглощение полностью отсутствует у молекул, поглощающий осциллятор которых перпендикулярен электрическому вектору возбуждающего света. В этом случае измеряют поляризационные спектры флуоресценции — зависимость степени поляризации флуоресценции объекта от длины волны возбуждающего света (поляризационные спектры по поглощению) или длины волны регистрации при фиксированной длине волны возбуждения (поляризационные спектры по испусканию).
Для количественной оценки поляризованной люминесценции используют степень поляризации (Р) люминесценции:
р = (1п - 1Х)/(1П + IJ,
где 1„ и 1_ — соответственно параллельная и перпендикулярная электрическому вектору возбуждающего света составляющие поляризованной люминесценции.
Степень анизотропии (г ) излучения определяется по формуле: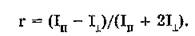
Использование этой характеристики для описания процессов поляризованной люминесценции более корректно, так как учитывает третью составляющую, перпендикулярную электрическому вектору возбуждающего света (т.е. в знаменателе уравнения для определения г находится величина, пропорциональная суммарному значению интенсивности).
Связь между величинами Риг отражает следующее уравнение:
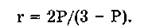
Анизотропия оптической среды проявляется как при испускании, так и при поглощении света. Для характеристики анизотропии поглощения света используют степень дихроизма d:
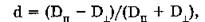
где Dn и D±—- оптические плотности, измеренные при облучении исследуемого образца линейно-поляризованным светом с колебаниями электрического вектора, параллельными и перпендикулярными главной оптической оси системы.
Перреном и Яблонским установлена связь между степенью поляризации (Р) флуоресценции и вязкостью среды (ц):
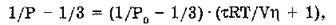
где Ро — предельное значение поляризации при Т/р —> 0; R — универсальная газовая постоянная; Т — абсолютная температура; V — объем моля флуоресцирующих молекул; т — время жизни возбужденного состояния. Этой формулой пользуются для расчета значений т.
Варьируя угол направления поляризации флуоресценции, устанавливают направление и степень ориентации молекул в анализируемом образце. Исследование поляризации флуоресценции позволяет изучать миграцию энергии в биосистемах, определять размеры биомакромолекул и время жизни их возбужденного состояния. Поляризацию флуоресценции определяют для того, чтобы судить о микровязкости мембраны.
Так, микровязкость мембраны изучают путем измерения Р для флуоресцентного зонда, введенного в исследуемую систему. Метод флуоресцентных зондов подробно рассмотрен в разделе 6.4.
В целом флуоресцентный метод анализа — один из безынерционных, высокочувствительных методов исследования, применяемых для качественного и количественного анализа смесей веществ, изучения механизма фотофизических и фотохимических процессов в биосистемах и конформационных свойств биомакромолекул.
Инфракрасная (ИК) спектроскопия занимается анализом молекулярных спектров, так как в ИК-области спектра (10 000—400 см'1) расположено большинство колебательных и вращательных спектров молекул. В ИК-диапазоне поглощается свет тех частот, которые равны частотам колебаний атомов в молекуле. ИК-спектр поглощения, возникающий в результате поглощения ИК-излучения при прохождении его через вещество, состоит из большого числа полос, причем часть полос поглощения обусловлена колебаниями отдельных атомных группировок, более или менее сохраняющих свои свойства в разных молекулах. Характеристики ИК-спектра (число полос поглощения, их положение, определяемое частотой v или длиной волны X, ширина и форма, интенсивность поглощения в максимумах) определяются структурой и химическим составом поглощающего вещества и зависят от его структурного состояния, а также температуры, давления и других факторов. Метод ИК-спектроскопии позволяет определять структуру молекул, их химический состав, моменты инерции, величины сил, действующих между атомами в молекуле, проводить качественный и количественный анализ смесей различных веществ.
Сущность одного из видов метода ИК-спектроскопии — ИК-дихроизма — заключается в измерении разности поглощения света, поляризованного вдоль оси ориентации и перпендикулярно этой оси ориентации молекулы. С помощью измерения ИК-дих- роизма можно установить, как направлены важнейшие группы белка (> С = О и -NH) по отношению к оси его макромолекулы. Необходимыми для определения частотами пептидной связи белков являются следующие: 3330 см'1 для валентного колебания - NH-группы (колебание ядер вдоль линии связи между атомами в молекуле), 1660 см'1 для валентного колебания >СО-группы и 1550 см'1 для деформационного колебания (колебания, обусловленные периодическими изменениями угла между связями отдельных атомов в молекуле ) NH-группы. Анализ смещения этих полос поглощения, изменения их ширины, формы, величины ПО-
глощения позволяет судить о состоянии вторичной структуры белков и полипептидов.
Дисперсия оптического вращения (оптической активности) — зависимость угла вращения (поворота) плоскости поляризации света (0), либо против часовой стрелки (ф0) и левовращающие (отрицательно вращающие — I, ф