Мембранное фосфорилирование и электронный транспорт
Важным и наиболее тщательно изученным процессом с участием молекулярных машин является образование аденозинтрифосфата (АТФ или АТР) — универсальной энергетической «разменной монеты» в биологических системах, и гидролиз АТР, который удовлетворяет энергетические потребности всех энергоакцепторных процессов, как, например, синтез низко- и высокомолекулярных соединений, создание неравновесного распределения ионов металла и других низкомолекулярных частиц внутри гетерогенных тканей и внутриклеточных структур, механическое движение, люминесценция и т.д.

Рис. 4.14. Структуры АТР и ADP. Схема синтеза и гидролиза АТР
На рис. 4.14 показаны структура ATP и ADP и схемы синтеза и гидролиза АТР, т. е. схемы реакции
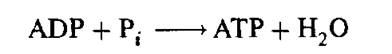
Как уже было ранее подчеркнуто, все биологические молекулярные машины являются машинами химическими.. Они осуществляют сопряжение между энергодонорными и энергоакцепторными химическими реакциями. В классической физической химии такое сопряжение требует присутствия общего интермедиата, участвующего в обеих реакциях.
Рассмотрим простейший пример с двумя мономолекулярними реакциями

Здесь К{ и К2 — константы равновесия реакций (4.21) и (4.22) соответственно.
Пусть реакция (4.21) — энергодонорная, а реакция (4.22) — энергоакцепторная. Детальный математический анализ этой ситуации можно найти в [31]. Здесь мы ограничимся объяснением, несколькими утверждениями и общим выводом.
Предположим, что наша система исходно находится в состоянии химического равновесия. Сдвиг равновесия энергоакцепторной реакции может быть вызван, например, повышением концентрации одного из реагентов, участвующих в энергодонорной реакции.
Этот механизм имеет чисто статистическую природу. Поскольку энергетическое сопряжение осуществляется благодаря изменениям в концентрации одного из участников, то за производство химической работы ответственна энтропийная часть изменения свободной энергии. Это энтропийный механизм непрямого энергетического сопряжения химических реакций. Рассмотрим его более подробно.Энергия субстрата первоначально сохраняется в форме потенциальной энергии химических связей молекулы. Энергия, освобождаемая в ходе энергодонорного процесса, распределяется по огромному числу тепловых степеней свободы общего резервуара, и не поступает
непосредственно для выполнения работы или повышения потенциальной энергии других реагентов. Элементарные акты энергодонорного и энергоакцепторного процессов могут быть разделены во времени и пространстве. Возникает вопрос: как может энергодонорная реакция стимулировать преодоление активационного барьера в ходе энергетически невыгодного химического превращения?
Следует отметить, что в случае энтропийного механизма непрямого сопряжения обе реакции могут быть как эндотермическими, так и экзотермическими. Для обеспечения сопряжения определяющую роль играет только знак изменения свободной энергии. Полная энергия системы может либо понижаться, либо возрастать, в зависимости от природы элементарного акта энергоакцепторной реакции. Энергия, необходимая для осуществления химического превращения либо забирается из окружения (термостаты) или ему передается. Изменения концентрации приводят только к изменению вероятности соответствующих актов. Молекулы, участвующие в элементарных актах с «энергоакцепторной стороны», выбирают «горячие» частицы из реагентов реакционной смеси. Они работают, на самом деле, как демон Максвелла (см. раздел 3.3). Химическая система освобождается от «информационного мусора», осуществляя превращение субстрата в продукт, и «демон Максвелла» может снова продолжать свою работу.
Однако количественный анализ показывает, что во всех случаях энтропийный механизм непрямого сопряжения характеризуется низкой эффективностью процесса.
Имеются две возможные меры непрямого энергетического сопряжения. Первая определяется как отношение числа результирующих элементарных актов энергоакцепторного процесса па к числу элементарных актов сопряженного энергодонорного процесса nd. Мы будем называть эту меру эффективностью сопряжения и обозначать ее 7 = njnd, Эту меру можно использовать независимо от метода инициирования энергодонорного процесса (добавление субстрата, изменение температуры, добавление катализатора к системе, первоначально находящейся в неравновесном состоянии). Вторая мера — это отношение па к числу добавленных молекул субстрата энергодонорной реакции nad, и обозначается как rj = njnai. Эту меру мы будем называть фактором конверсии. Эта мера особенно полезна в случае стационарного состояния открытых систем, с непрерывным добавлением субстрата. Анализ показывает [31,41], что эффективность сопряжения достаточно высока (приближается к 1), только если энергоакцепторная реакция термодинамически выгодна, т. е. может идти без какого-либо энергодонорного процесса. Чтобы достичь больших значений фактора конверсии т/, мы должны иметь огромный избыток концентрации субстрата. Так, например, в случае синтеза АТР, чтобы образовать одну молекулу С, нужно добавить больше чем 106 молекул В. Оба эти условия иногда не выполняются в биоэнергетических процессах.Вернемся теперь к синтезу АТР. Подавляющая часть молекул АТР (около 85 %) в животных бактериальных и растительных клетках синтезируются в мембранных внутриклеточных структурах (мембранное фосфорилирование). В аэробных организмах непосредственными источниками энергии (энергодонорные процессы) являются определенные стадии окисления пищи. В растениях и фотосинтезирующих бактериях первичными источниками являются, конечно, кванты света, энергия которых, после возбуждения хлорофилла, превращается в энергию в окислительно-восстановительных цепях электронного транспорта (ЦЭТ) в тилакоидных мембранах хлоропластов.
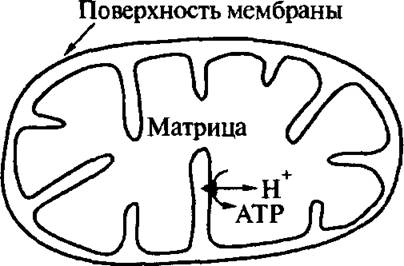
Рис. 4.15. Схематическое изображение митохондрии
На рис. 4.15 и 4.16 изображены схемы поперечного сечения митохондрии и ее ЦЭТ, локализованной во внутренней мембране митохондрий. Митохондрии являются внутриклеточными органеллами. Их внутренние мембраны содержат «электронные переносчики» и ферменты, катализирующие синтез АТР из аденозинди- фосфата (ADP) и неорганического фосфата Р; (см. рис. 4.14). Каждая
ЦЭТ содержит три идентичных ATP-синтезирующих ферментных комплекса (АТРсинтаза). Этот же фермент катализирует и гидролиз АТР. В этом случае его называют АТРазой. Митохондрия представляет собой продолговатую, эллипсоидальную частицу (длинная ось — примерно 3 микрона, короткая ось — примерно 1 микрон). Она окружена двумя
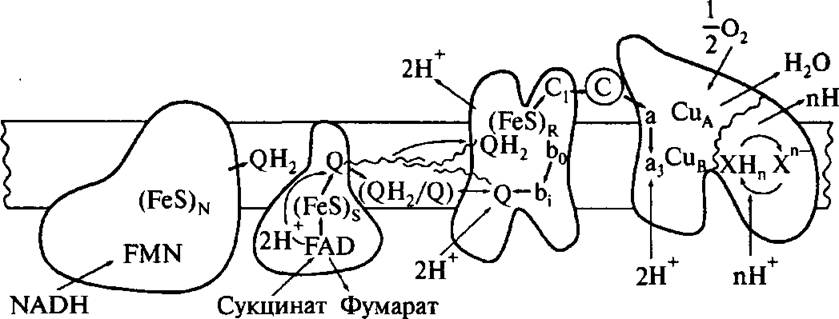
Рис. 4.16. Схема электронного и протонного переноса во внутренней мембране митохондрии
мембранами: гладкой наружной мембраной и внутренней мембраной с кристами, образующими довольно сложный рельеф. Внутренняя мембрана одной митохондрии содержит около 1 500 ЦЭТ. Каждая ЦЭТ может быть разделена на несколько частей, называемыми обычно «комплексами Грина». Два субстрата ЦЭТ, NADH и сукцинат, взаимодействуют с ЦЭТ через первый и второй комплексы Грина, соответственно. Эти комплексы содержат флавопротеины (FP). Их активными центрами являются флавинмононуклеотид (FMN, комплекс 1), флави- наден инд и нуклеотид (FAD, комплекс 2), а также железосерные белки (Fe-S-белки), обозначаемые N (комплекс 1) и S (комплекс 2). Два пути электронного транспорта от NADH и сукцината соединяются в «уби- хинонном котле». Убихинон (UQ), низкомолекулярный электронный переносчик (называемый также «коэнзимом Q10» или просто коэнзимом Q), переносят электроны между центрами, локализованными на разных сторонах внутренней мембраны митохондрии, одновременно обеспечивая перенос протонов поперек мембраны (рис.
4.16).ЦЭТ митохондрий содержит три центра сопряжения, локализованные в первом, третьем и четвертом комплексах Грина (см. рис. 4.15 и 4.16). В первом комплексе центром сопряжения служит Fe-S белок N-2, во втором — Fe-S белок «Риске», а в четвертом — цитохром- С-оксидаза. Прохождение электрона через центр сопряжения сопровождается понижением его энергии, что впоследствии используется соответствующей АТРсинтазой для синтеза АТР Перенос одного электрона на центр сопряжения от предыдущего переносчика означает восстановление первого и окисление последнего.
Энергодонорная и энергоакцепторная реакции могут быть разобщены весьма слабыми воздействиями: нагреванце до 50° С, повторным замораживанием и оттаиванием, старением. В разобщенных митохондриях энергия, освобождаемая при окислении NADH или сукцината диссипирует, и АТР не синтезируется. В разобщенном состоянии, когда процесс электронного транспорта не связан с синтезом АТР, его скорость резко возрастает. Образование АТР не идет также и в сопряженных митохондриях в отсутствие, например, субстратов фосфорилирования (ADP или Pf), но в этом случае скорость электронного переноса понижается. Таким образом, не только энергодонорная реакция необходима для реализации энергоакцепторной, но и, наоборот, энергоакцепторная реакция необходима для реализации энергодонорной! Это совершенно необычная ситуация для классических химических реакций. Это напоминает поведение двух соединенных шестеренок, которые не могут вращаться друг без друга. Как будет показано далее, для процессов трансформации энергии в биологических системах эта механическая аналогия вполне реальна.
Детальные кинетические исследования, проведенные в лаборатории Чанса [54,55], показали, что отсутствие сопряжения ускоряет перенос электрона через центр сопряжения. Эти стадии и определяют скорости электронного транспорта в митохондриях.
На рис. 4.17 представлена схема процесса сопряжения. В этом случае энергодонорной реакцией является перенос электрона от первоначального восстановленного переносчика (DR) на следующий переносчик ЦЭТ (Ад), который перед электронным переносом был окислен.
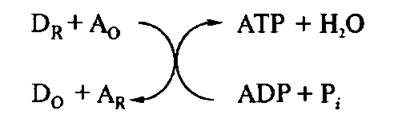
Рис. 4.17. Схема процесса сопряжения в центре сопряжения
Эта схема работает, как для центров сопряжения ЦЭТ митохондрий, так и для ЦЭТ хлоропластов и фотосинтетических бактерий.
Ферменты, осуществляющие синтез АТР также идентичны во всех этих системах.
В 1962 году Шен и Шен (КНР) выполнили очень важный эксперимент, изучая индуцируемое светом мембранное фосфорилирование в хлоропластах высших растений [56]. Освещение хлоропластов в отсутствие субстратов фосфорилирования (ADP и PJ приводит к образованию некоторого энергизованного соединения или состояния ж, которое способно обеспечить синтез АТР в темноте, после добавки ADP и Р:. Такие же эксперименты с теми же результатами были независимо (никто не читает китайских журналов) выполнены в США [57]. ж не может быть образовано при температурах ниже -13° С, хотя индуцированный электронный транспорт продолжается и при температурах до -30° С. В отсутствие АТР и фосфата время жизни ж в темноте равно около 0,5-1 с при 20° С и 45 с при 0° С. Образование ж сопровождается конформационными изменениями мембраны. Эти изменения, вероятно, являются вторичными процессами. Если ж образуется во время освещения, то в темноте после добавления субстрата фосфорилирования может быть синтезировано до 30 молекул АТР в каждой ЦЭТ [56-58].
Таким образом, ясно, что в схему, показанную на рис. 4.17, мы должны включить ж, которую обычно называют энергизованным интермедиатом или первичным макроэргом (рис. 4.18).
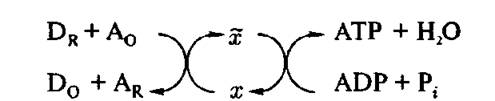
Рис. 4.18. Схема процесса сопряжения в центре сопряжения с участием первичного макроэрга
Все гипотезы о возможных механизмах мембранного фосфорилирования отличаются друг от друга предположениями о природе ж. Первая концепция была предложена Е. Слейтером в 1953 году. По Слейтеру, х — это некоторое «энергизованное химическое соединение». После бесчисленных напрасных попыток обнаружить х, в 1961 году Митчелл и Вильямс независимо высказали идею о том, что образование АТР может происходить под действием протонов без какого-либо химического интермедиата [50, 60]. Эта очень плодотворная идея дала сильный толчок развитию биоэнергетики за последние 35 лет. Между точками зрения Митчелла и Вильямса о роли протонов как макроэргов была, однако, большая разница^ Эта разница сводилась не только к языку, использованному обоими авторами, но и имела принципиальное значение для истинного понимания механизма мембранного фосфорилирования. В своей «хемоосмотической» гипотезе Митчелл связывает протондвижущую силу с трансмембранной разницей электрохимических потенциалов, возникающих в водных фазах, разделенных мембраной. Включение в рассмотрение обширных водных фаз с необходимостью требует диссипации энергии, освобождаемой в ходе энергодонорного процесса (электронный транспорт) по всему окружающему пространству. Отвергая эту точку зрения, Вильямс утверждает: «Протоны в мембранах, а не осмотический трансмембранный градиент протонов требуются для обеспечения синтеза АТР». Он говорит также об энергиях локальных зарядов, генерируемых при отделении протонов от электронов соответствующими реакциями.
В настоящее время ясно (мне, по крайней мере), что Вильямс был прав, а Митчелл ошибался.
Более того, эти два ученых отвечали, по существу, на разные вопросы. Подход Митчелла строго следовал обычной термодинамике. Он ответил на следующий вопрос: может ли в принципе перенос протонов вдоль градиента их электрохимического потенциала Д/хн+ обеспечить энергоакцепторный процесс синтеза АТР? Ответ, конечно, да. Это термодинамически возможно, если Д^н+ удовлетворяет условию:

где Д6? — возрастание свободной энергии Гиббса, требуемое для образования АТР из ADP и Р-. Митчелл — автор многих блестящих публикаций, относящихся к проблемам переноса ионов через биологические мембраны. Его статьи фактически создали новую главу в биохимии живой клетки. Он был заслуженно удостоен Нобелевской премии. Однако в своих статьях, касающихся хемоосмотической концепции, он ни разу не ответил и даже ни разу не задал единственный важный вопрос в этой проблеме: как может энергодонорная реакция переноса протона через мембрану быть прочно сопряжена с энергоакцепторной реакцией образования АТР? Подход Митчелла в сущности сводит проблему к закону действующих масс, т. е. к схеме реакций, представленных в уравнениях (4.20) и (4.21), которая никогда не может привести к значительной эффективности.
С другой стороны, по Вильямсу, первичный макроэрг представляет собой локальный зарад, образованный при отрыве протона от электрона в ходе соответствующей реакции. Возникновение такого протона на самом деле представляет собой образование неравновесного состояния, определенного локального центра. Это согласуется с концепцией МакКлэйра о том, что сохраняемая энергия может выполнить работу только в ходе переноса к другой форме сохраняемой энергии.
Детальный анализ различи^ концепций Митчелла и Вильямса можно найти в монографиях [31,41]. Рассмотрим теперь образование таких неравновесных состояний в электрон-транспортной цепи митохондрий и в ферменте АТРсинтазы.
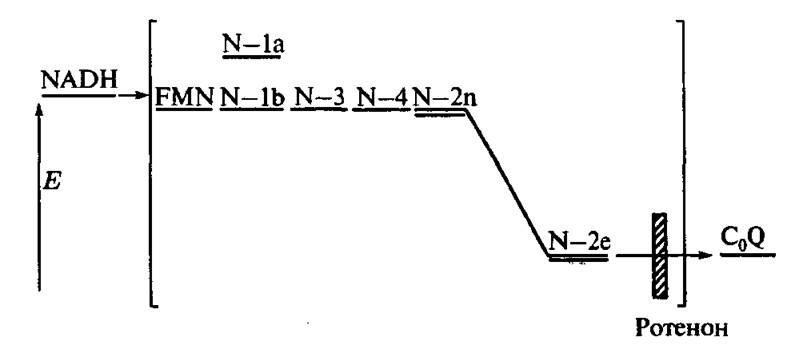
Рис. 4.19. Схема ЦЭТ вблизи первого пункта сопряжения и положения уровней энергии восстановленных электронных переносчиков
На рис. 4.19 представлена ЦЭТ в первом комплексе Грина и даны положения уровней энергии электрона в восстановленных состояниях соответствующих электронных переносчиков. Можно видеть, что единственное существенное уменьшение энергии электронов имеет место при переходе от железосерного,центра N-2 к убихинону. Было доказано, что в первом пункте сопряжения митохондрий роль «переносчика-трансформатора» играет N-2 центр [61].
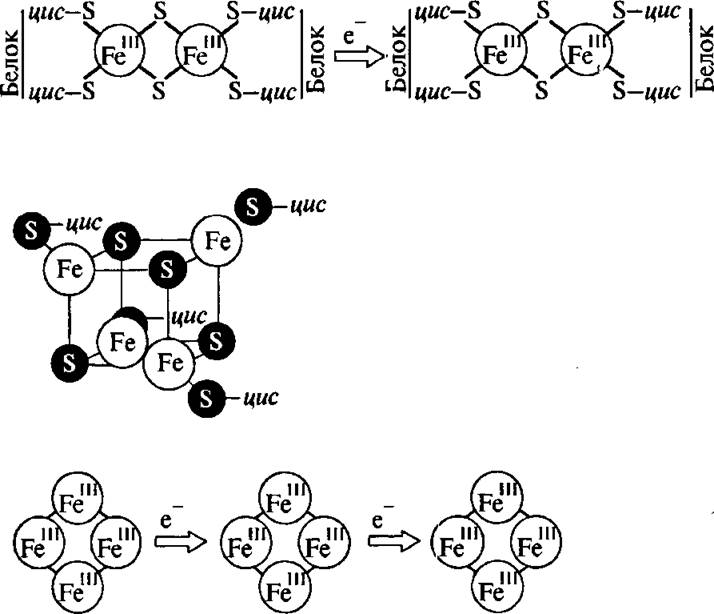
Рис. 4.20. Структуры активных центров биядерных и тетроядерных железосерных белков
На рис. 4.20 изображены структуры активных центров в биядерных и тетроядерных железосерных белках. 5)ти центры, как правило, содержат два или четыре атома железа. Каждый атом железа находится в тетраэдрическом окружении серных ли гандов: два цистеиновых остатка соединяют железо с белком, а два атома неорганической серы образуют мостики между атомами железа. Железосерные центры являются одноэлектронными переносчиками. В окисленном, основном, состоянии центр диамагнитен. Одноэлектронное восстановление превращает его в парамагнитный центр, который может быть зарегистрирован методом ЭПР.
ЦЭТ митохондрий может находиться в нескольких состояниях [54,55]. Для нас важны сопряженные состояния 3 и 4. В состоянии 3 имеется избыток субстратов фосфорилирования и образование АТР не является лимитирующей стадией всего процесса. В состоянии 4
концентрация ADP очень мала и лимитирующей стадией скорости всего процесса сопряжения становится акт синтеза АТР.
Наиболее информативным методом изучения окисления и кон- формационных состояний железосерных белков является ЭПР. Этот метод был применен для регистрации неравновесного состояния центра N-2 в первом комплексе Грина и его релаксации к равновесию.
Для биядерных центров действует модель обменного анти- ферромагнитного спинового взаимодействия. Согласно этой модели, основное состояние восстано
вленного центра имеет суммарный спин S = х!г. Неспаренный электрон делокализован между двумя атомами железа с обменным интегралом J < 0. Первый возбужденный спиновый уровень (5 = 3/>) лежит на А = 3|J| над основным уровнем. В качестве первого приближения можно считать, что это справедливо и для тетроядерных центров.
Спин-решеточная магнитная релаксация (не путать с конформационной релаксацией!) железосерных центров осуществляется по двухфонному механизму Орбаха (рис. 4.21).
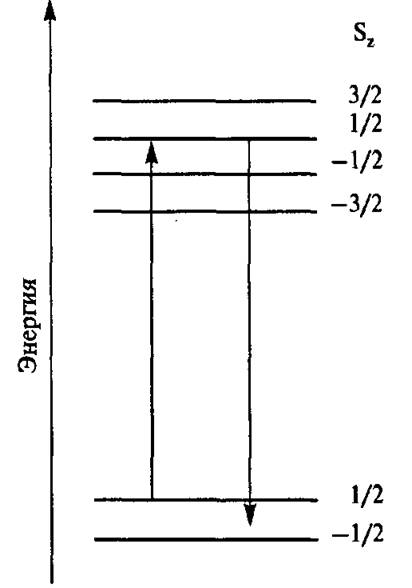
Рис. 4.21. Механизм Орбаха спин-решеточной релаксации
Чтобы получить железосерный центр в неравновесном состоянии, был использован описанный выше метод низкотемпературного восстановления термолизованными электронами. Повышение температуры приводит к усилению внутримолекулярной подвижности и релаксации центра к равновесному состоянию.
На рис. 4.22 представлен центр ЭПР митохондрий сердца быка при 20 К. Эта температура соответствует максимальной интенсивности сигналов ЭПР железосерных центров — см. ниже). Митохондрии были в восстановленном состоянии. Восстановителями служили эндогенные
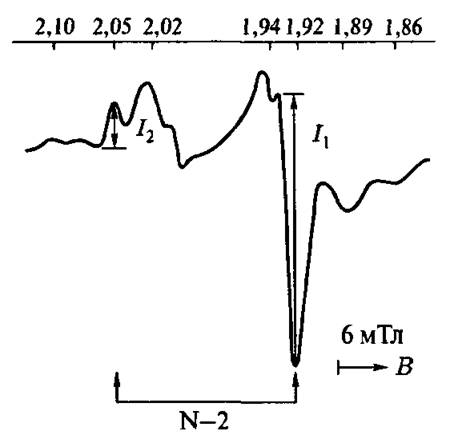
Рис. 4.22. Механизм Орбаха спин-решеточной релаксации
субстраты при комнатной температуре, так что все парамагнитные центры успевали релаксировать. Две компоненты, обозначенные стрелками, принадлежат центру N-2. Их положения (//-факторы) в неравновесном состоянии не изменяются. Однако температурные зависимости этих компонент чрезвычайно чувствительны к ничтожным структурным изменениям.
Поэтому для изучения неравновесных состояний железосерных центров, появляющихся в процессе электронного переноса, была использована так называемая техника кривых 1(Т) — «кривые зависимости интенсивности от температуры». На рис. 4.23 показана типичная кривая ЦТ) железосерного центра (в данном случае — восстановленного ферредоксина). В низкотемпературной области (1) интенсивность сигнала уменьшается с уменьшением температуры, благодаря эффектам насыщения микроволновой мощностью. В срединной части (2) выполняется закон Кюри (I = const/T). Отклонение от закона Кюри в высокотемпературной части (3), обусловлено уширением линии, являющейся следствием уменьшения времени спин-решеточной релаксации. Положение кривой ЦТ) (особенно ее максимума) на оси Т является «отпечатком пальцев» структуры центра. Согласно механизму Орбаха, магнитная релаксация (и поэтому микроволновое насыщение) центра сигнала ЭПР, определяется значением обменного интеграла J (см. выше). Это значение, в свою очередь, зависит главным образом от расстояния между атомами железа и от значений S-Fe-S углов в железосерном центре (рис. 4.20).
На рис. 4.24 представлены кривые ЦТ) восстановленного центра N-2 митохондрий сердца быка, зарегистрированные в разных условиях. Наиболее интересные особенности этих данных могут быть вкратце сформулированы следующим образом. Кривая ЦТ) центра N-2
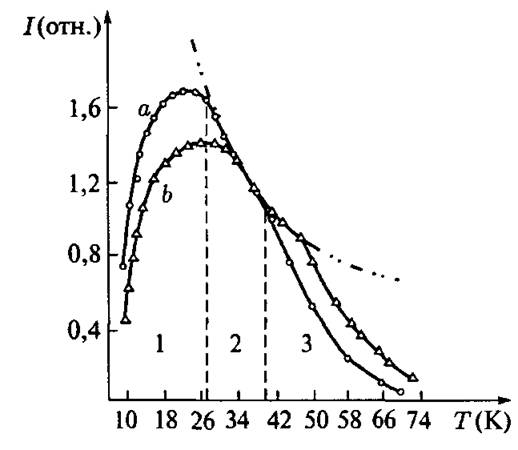
Рис. 4.23. Кривая 1(Т) сигнала ЭПР (компонента gz) восстановленного растворимого фер- редоксина гороха в равновесном состоянии: a — исходный препарат, б — слегка конформационно измененный после добавки 0,8 М NaCl
восстановленного NADH сопряженных митохондрий в состоянии 4, совпадает с кривой J(T) неравновесного центра N-2 в разобщенных митохондриях, полученных восстановлением термолизованными электронами в замкнутой матрице.
Рис. 4.24. Кривые 1(Т) восстановленного N-2 центра митохондрий сердца быка: (1о) — разобщенные митохондрии, восстановленные термолизованными электронами при 77 К; (2*) — тот же препарат после релаксации при 210 К; (1а) — активные фос- форилирующие митохондрии в состоянии 4. Недостаточно ADP, и скорость электронного транспорта определяется скоростью фосфорилирования ADP; (2 ж) — тот же препарат после разобщения методом оттаивания и замораживания
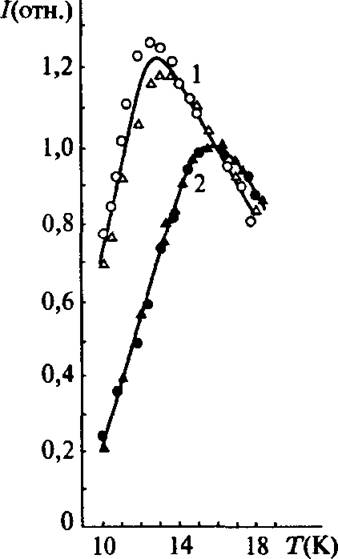
В третьем состоянии кривая ЦТ) совпадает с кривой равновесного центра N-2 в разобщенных митохондриях. Такая же кривая ЦТ) может быть получена и у сопряженных митохондрий, если заблокировать ротеноном перенос электрона между N-2 и UQ (рйс. 4.19).
Чтобы понять значение этих данных, полезно рассмотреть изменения энергии электрона на его пути через первый комплекс Грина (4.19). Перенос электрона через ряд переносчиков от NADH до N -2 не сопровождается изменением его энергии. Следующий акт, восстановление UQ, может произойти только после понижения энергии неравновесного восстановленного центра N-2 в ходе конформационной релаксации. Анализ лимитирующих стадий дыхательного электронного транспорта, проведенный во многих лабораториях с различными субстратами (наиболее важные результаты были получены в лаборатории Б. Чанса [41,61]), показали, что в четвертом дыхательном состоянии перенос электрона от NADH к UQ замедляется на 30-50 мс по сравнению с третьим дыхательным состоянием (избыток ADP).
Таким образом, перенос электрона от NADH к N-2 не является лимитирующей стадией процесса электронного переноса от NADH к UQ. Эксперименты с ротеноном показали, что во время прямого электронного переноса конечное состояние центра N-2 равновесно. Это значит, что в этом случае неравновесное восстановленное состояние N-2, образованное после присоединения электрона к равновесному окисленному центру, переходит в равновесное состояние. Более того, это также значит, что перенос электрона от N-2 (после релаксации) к следующему акцептору (UQ) тоже не является лимитирующей стадией. Замедление электронного транспорта в четвертом дыхательном состоянии вызвано, таким образом, возрастанием времени перехода восстановленного неравновесного центра N-2 в равновесное состояние. Это может быть связано либо с временем жизни центра N-2 в неравновесном состоянии, либо с длительностью конформационной релаксации. Неравновесный восстановленный центр N-2 был зарегистрирован в митохондриях в четвертом дыхательном состоянии. Поэтому можно сделать вывод, что лимитирующей стадией является первая из перечисленных выше, т.е. время жизни N-2 в неравновесном восстановленном состоянии. Следует напомнить, что в четвертом дыхательном состоянии электронный транспорт происходит гораздо медленнее, чем в третьем, и что в нативных прочно сопряженных митохондриях синтез АТР практически прекращается [62]. Отсюда следует, что в этих условиях перенос электрона через пункт сопряжения и, следовательно, релаксация центра N-2 в равновесное состояние не может происходить без фосфорилирования АЕ)Р.
Энергия, освобождаемая при окислении NADH, сохраняется в конформационно неравновесной форме N-2 и может быть затем использован для фосфорилирования ADP АТРсинтазой.
4.9.
Еще по теме Мембранное фосфорилирование и электронный транспорт:
- Цикл трикарбоновых кислот, транспорт электронов
- Статья 62. Реестр участников электронного аукциона, получивших аккредитацию на электронной площадке
- Статья 61. Аккредитация участников электронного аукциона на электронной площадке
- Выбор операторов электронных площадок в целях проведения открытых аукционов в электронной форме на право заключения государственных и муниципальных контрактов
- Плазматическая мембрана
- Мембраны первого типа
- Проницаемость природных мембран
- Статья 59. Аукцион в электронной форме (электронный аукцион)
- Мембраны третьего типа
- Мембраны второго типа
- Мембраны четвертого типа
- Цитоскелет (мембранный каркас)
- 1.2.1. Классификация, структура и функции мембранных белков
- Роль жирных кислот мембранных липидов в холодоустойчивости растений
- Особенности структуры и функций мембранных рецепторов
- Нарушение структурно-функциональных свойств клеточных мембран