Элементарный акт функционирования АТРсинтазы
Научная общественность встретила без энтузиазма первую статью Митчелла [59], посвященнуго хемоосмотической гипотезе. Только после публикации в 1966 блестящих экспериментов по образованию АТР после быстрого скачка pH [63] хемоосмотическая гипотеза была почти единодушно принята специалистами.
Эти эксперименты были поставлены Ягендорфом и Юрайбом с адаптированными к темноте хлоропластами. Субстраты фосфорилирования были добавлены в темноте, и образец инкубировали также в темноте при низких значениях pH до тех пор, пока значения внутреннего pH (pHin) и внешнего (pH0Ut) не выравнивались. Зачем (pH0Ut) быстро (~ 0,5 с) повышали до 9-9,5. В темновых; хлоропластах нет электронного переноса и синтеза АТР. Тем не менее, щелочной скачок сопровождается образованием АТР при комнатной темпералуре в течение нескольких секунд, а при 4° С — целую минуту. Ягендорф и Юрайб в этих опытах создали искусственно в темноте типич ную «Митчелловскую» ситуацию: высокое значение pH0Ut и низкое — pHjn. Оказалось, что градиента pH вполне достаточно, чтобы обеспечить условия, необходимые для синтеза АТР. После того, как множество лабораторий во всем мире воспроизвели эти результаты, после того, как было показано, что действие различных ингибиторов на естественное мембранное фосфорилирование идентично их действию на фосфорилирование, инициируемое скачком pH, гипотеза Митчелла съдла общепринятой. Все были счастливы.Только одно заемное облако портило этот прекрасный ландшафт. Митохондрия — это вывернутый наизнанку тилакоид хлоропластов.
Согласно концепции Митчелла синтез АТР в условиях, аналогичных опытам Ягендорфа и Юрайба, требует не повышения, а скачкообразного понижения pH. Такие опыты были проведены во многих лабораториях сразу после публикации работы Ягендорфа и Юрайба, но безуспешно.
Насколько я знаю, появилась только одна статья, посвященная pH индуцированному образованию АТР в митохондриях [64]. Эти эксперименты были выполнены в лаборатории Митчелла следующим образом. Митохондрии печени крысы инкубировали при pH 7,0 в присутствие ADP и Р-. Затем pH повышали до 9,0, а через некоторое время снова понижали до 4,4. В результате этой процедуры содержание АТР в образце в расчете на одну ЦЭТ повышалось приблизительно на три молекулы. Предварительное повышение pH до 9,0 было необходимо, по мнению авторюв, для достижения достаточно большой величины ДрН после после дующего его понижения. Меньшее значение ДрН было бы не эффект ивно (не хватало энергии), а при pH ниже 4,4 митохондрии дезактивируются.В нашей лаборатории были пост авлены специальные эксперименты (все ссылки на эти работы и на опыты, выполненные затем в других лабораториях можно найти в [1, 31,4Ї,61]). Было показано, что в экспериментах [64] дополнительное количество АТР образуется на стадии предварительного повышения: pH от 7,0 до 9,0, а не на стадии последующего понижения pH. Митохон дрии, таким образом, требуют тот же знак ДрН, что и хлоропласты.
Первые эксперименты были поставлены нами с сопряженными и разобщенными митохондриями. Основные результаты: сопряженные митохондрии способны осуществлять количественный синтез АТР в ответ на быстрое повышение pH. В расчете на одну ЦЭТ один pH скачок приводит к образованию от двух до пяти (в зависимости от препарата) молекул АТР. Обязательным условием является пересечение во время скачка значения pH ~ 8,2. Те же результаты были получены с разобщенными митохондриями, не способными к окислительному фосфорилированию. На рис. 4.25 о приведена экспериментальная зависимость выхода АТР при быстром повышении pH в суспензии разобщенных митохондрий печени крысы от конечного значения pH при фиксированном начальном (кривая 1) и от начального значения pH при фиксированном конечном (кривая 2). Эти зависимости соответ-
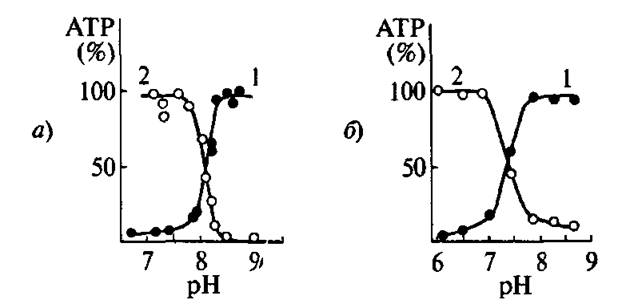
Рис.
4.25. Зависимость выхода /АТР от конечного значения pH при фиксированном начальном (1) и от начального значения pH при фиксированном конечном. а) Разобщенные митохондрии печени крысы; б) фрагменты стенок мембраны из Staphylococcus aureusствуют кривым титрован мя кислотных групп с рК « 8,2. Индуцируемое скачком pH образование АТР не нуждается в функционировании ЦЭТ, полностью блокируется ингибиторами АТРсинтазы. Синтез АТР не наблюдается в отсутствии субстратов фосфорилирования. При избытке субстратов фосфорил ирования можно таким способом получить с одним и тем же образцом множество молекул АТР в расчете на одну ЦЭТ, меняя попеременно pH, например, от 7,5 до 8,5 и обратно. АТР образуется только на стадии повышения pH.
Если ADP и pj добавляют после «правильного» скачка pH, то выход АТР зависит от промежутка времени между скачком pH и добавкой. Для митохондрий время релаксации Ту2, т.е. интервал времени, после которого выход АТР уменьшается наполовину, составляет при комнатной температуре около 10 с. Изолированные АТРсинтазы (см. ниже) релаксируют быстрее.
Образование АТР в ответ на быстрый скачок pH наблюдали также с мембран связанными АТРазами без замкнутых везикул (рис. 4.25 б). Для осуществления элементарного акта синтеза АТР после скачка pH не нужны ни замкнутые везикулы, ни даже фрагменты мембран. Результаты, аналогичные описанным выше, были получены также с изолированными растворимыми компонентами АТРаз животных, бактериальных и растительных клеток. Этот эффект является общим свойством не только АТРаз из трансформирующих энергию мембран, но и других
АТРаз, нормальное функционирование которых не сопровождается образованием АТР, например АТРаз миозина, нитрогеназы и др.
В рамках описанного выше релаксационного механизма ферментативного катализа энергодонорной реакцией, сопряженной с энергоакцепторной стадией синтеза АТР, является акт диссоциации определенной кислотной группы (или групп) в каталитической части АТРсинтазы.
Рассмотрим теперь структуру АТРсинтазы и попробуем ответить на главный вопрос: какая стадия процесса образования АТР в водной фазе является энергоакцепторной?
На рис. 4.26 представлена упрощенна^ схема АТРсинтазы. Структура состоит из двух основных частей: гидрофильного, экспонированного в водную фазу комплекса F! (фактор сопряжения) и гидрофобного фактора Fo, связанного с мембраной.
Fo выполняет роль специфического протонного канала, с помощью которого ионы водорода пересекают імембрану. В отсутствие субстратов фосфорилирования утечка протонов из водной фазы через комплекс Fo затруднена. Удаление фактора F,’ делает сопрягающую мембрану прозрачной для ионов водорода: появляется много сквозных дырок. Протоны в этом случае свободно проходят через мембрану, и их концентрации по обе стороны выравниваются. ;
Фактор сопряжения Fj представляет собой белковый комплекс, содержащий девять субъединиц: три одинаковых о субъединиц, три одинаковых р субъединиц и по одной 7, б и є субъединиц. Все а и р субъединицы расположены вокруг продолговатой 7 субъединицы (рис. 4.26). Фактор Fj имеет вращательную симметрию третьего порядка. Поворот вокруг оси, параллельной 7 субъединице, приводит к смещению каждой из а и р субъединиц в положение другой аналогичной субъединицы. 7 субъединица длиной 9 нм слегка изогнута и выступает на 3 нм из гексамера ЗаЗр (рис. 4.26). Субъединиц^ 7 и маленькая субъединица д играют роль мостиков, соединяющих части Fo и Ft АТРсинтазы. Небольшая субъединица є является регулятором ферментативной активности. В работах лаборатории Уокера можно найти детальное описание компонентов и структуры АТРсинтазы митохондрий.
В 1973-1974 годах две независимые группы ученых из лаборатории Бойера [65,66] и Слэйтера [67-69] предположили, что энергия, посту-
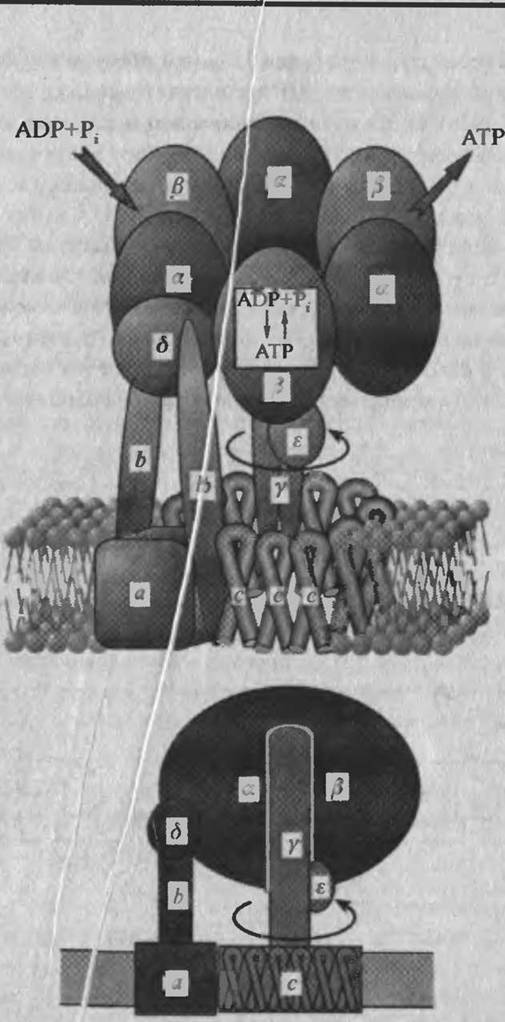
Рис 4 26 Пространственная структура АТРсинтазы
пающая в АТРсинтазу, расходуется главным образом на освобождение прочно связанной молекулы АТР из каталитического центра Fj.
Эта идея явилась основой на пути к по ниманию механизма мембранного фосфорилирования.По данным рентгеноструктурноіго анализа, молекулы АТР и ADP связываются между субъединицами а и /3 [73]. Синтез и гидролиз АТР катализируют только ft субъединицы. Молекулы АТР и ADP, связанные с а субъединицами, удерживаются так прочно, что во вре- мя работы фермента практически не обмениваются с молекулам АТР и ADP окружающей среды. Вероятно, эти прочно связанные молекулы не участвуют в синтезе АТР, а выполняют только регуляторные функции. На рис. 4.27 схематически показана последовательность событий
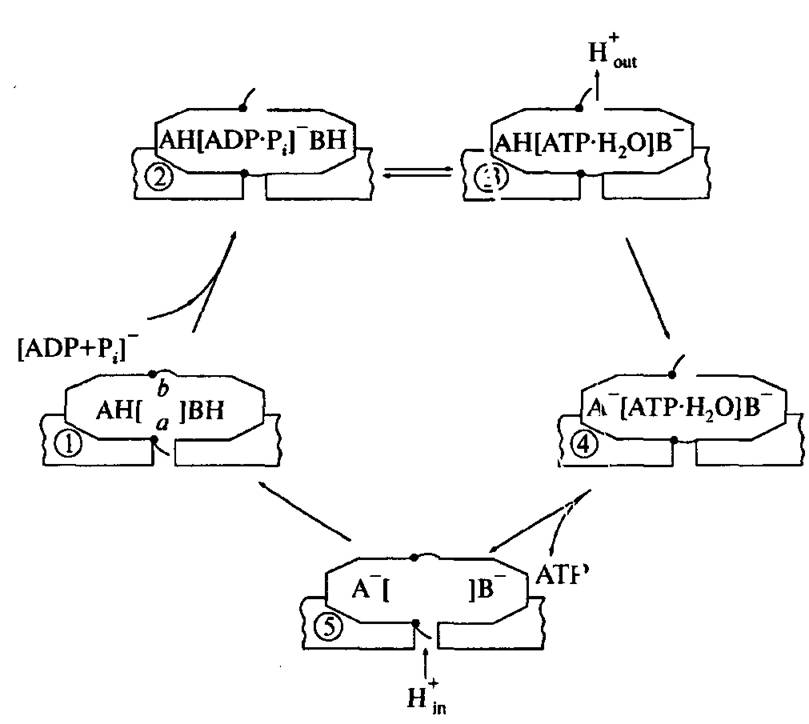
Рис. 4.27. Схема энергозависимых изменения схютояний в ходе синтеза АТР (объяснения в тексте *)
в ходе образования АТР. Первая стадия этого процесса — связывание ADP и р(. активным центром р субъединицы, находящейся в «открытом» конформационном состоянии 1.
В водной среде константа равновесия реакции
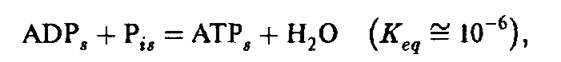
образование АТР энергетически невыгодно и равновесие реакции смещено влево. Здесь и далее нижние индексы s и Ъ означают, соответственно, «в водном растворе» и «связанный с мембраной». Присоединение субстрата (субстратов) фосфорилирования приводит, как всегда после связывания низкомолекулярного лиганда белковой глобулой, к возникновению неравновесного состояния Р субъединицы и ее релаксации к «закрытому» состоянию 2. По-видимому, первое предположение о конформационных изменениях ферментов при мембранном фосфорилировании было опубликовано в 1972 году [70]. Согласно измерениям Бойера, воспроизведенных во многих лабораториях, отношение ATP6/ADP6 = 1. Это значит, что AG = 0 и образование АТР происходит практически бесплатно. Вновь образованная молекула АТР прочно связана с Р субъединицей, и для ее освобождения в водную среду требуется затратить энергию на конформационную релаксацию субъединицы к «открытому» состоянию (стадия 3). Согласно описанным выше экспериментам со скачкообразными повышениями pH, эта последняя стадия начинается с диссоциации некоторых кислотных групп белковой глобулы.
Еще по теме Элементарный акт функционирования АТРсинтазы:
- 2.Понятие элементарной функции. Основные элементарные функции и их графики (постоянная, степенная, показательная, логарифмическая).
- Правовой акт государственных органов должен отвечать следующим требованиям: правовой акт не должен противоречить
- Глава 17. ПРАВОВИЙ АКТ. НОРМАТИВНО-ПРАВОВИЙ АКТ. МІЖНАРОДНИЙ ДОГОВІР
- § 13. Підзаконний нормативний акт державного підприємства, установи, організації. Підзаконний нормативний акт комерційної організації
- Элементарные преобразования.
- Пространство элементарных событий
- § 12. Підзаконний нормативний акт органу місцевого самоврядування. Підзаконний нормативний акт місцевого органу виконавчої влади
- § 31. Сведение к элементарным выражениям
- Интегрирование элементарных дробей.
- § 32. Элементарные выражения силлогистики
- Модель элементарной частицы.
- Элементарные преобразования систем.
- 3.4.2. Элементарный и полный импульс силы
- Акт толкования права.
- Элементарные преобразования деревьев
- 7. Общая характеристика низшего уровня элементарной сенсорной психики.
- 9. Общая характеристика высшего уровня элементарной сенсорной психики.
- §11. Відомчий акт
- Непрерывность некоторых элементарных функций.