Конечные продукты неферментативного гликирования белков и липидов
Наиболее распространенной гипотезой развития сосудистых осложнений сахарного диабета на сегодняшний день является неферментативное гликирование белковых аминогрупп, приводящее к образованию вначале легко обратимых шиффовых оснований в ходе реакции Мэйларда с последующим их превращением в более стабильные продукты Амадори, которые могут диссоциировать с высвобождением свободной глюкозы и молекулы протеина, но могут также, через стадию 3-деоксиглюкозона, медленно превращаться в стабильные и неподдающиеся расщеплению КПНГ (так называемые «коричневые продукты») [375, 376, 1676] (рис.
20).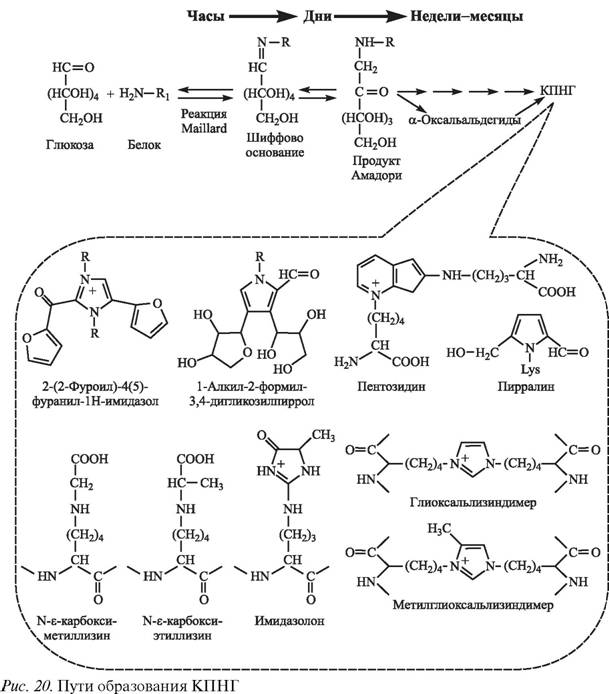
Формирование КПНГ может индуцироваться и другими α-оксальальдегидами — глиоксалем и метилглиоксалем, образующимися из трехуглеродных интермедиатов гликолиза — глицеральдегид-3-фосфата и дигидроацетонфосфата [307, 1409]. Все эти α-оксальальдегиды — 3-деоксиглюкозон, глиоксаль и метилглиоксаль — являются высокоактивными соединениями, обеспечивающими окислительное гликирование белков, энзимов и других важных клеточных компонентов [306, 964]. Они реагируют с лизиновыми и аргининовыми остатками протеинов, формируя специфические КПНГ, такие как пентозидин, пирралин, глиоксальлизиндимер, карбоксиметиллизин, карбоксиэтиллизин, имидазолон, метилглиоксальлизиндимер (см. рис. 20). Однажды сформировавшись, КПНГ остаются стабильными и необратимыми, аккумулируясь на белках сосудистой стенки и увеличиваясь в содержании в соответствии с концентрацией глюкозы [375]. В настоящее время общепризнанно, что КПНГ являются не только необратимыми продуктами, но могут быть и активными интермедиатами в межбелковых соединениях и генерации активных форм кислорода [1214, 1473]. Так, показано, что в результате гликирования в белках образуются ферментоподобные активные центры, которые представляют собой перекрестно связанные катион-радикалы шиффовых оснований протеинов и имитируют характеристики металлсодержащих окислительных систем, катализирующих генерацию АКМ [1815].
Повреждение белков в результате гликозилирования может служить фактором риска развития негативных процессов при диабете, в частности, атеросклероза, диабетической нефропатии и катаракты [1573]. В нормальных условиях у человека гликозилировано 1,3—2 % лизиновых остатков в липопротеинах низкой плотности, при диабете это значение возрастает в 2—3 раза [881]. В первую очередь гликированию подвергаются апо В [375] и фосфолипидные компоненты ЛНП [386], что приводит к изменениям их клиренса и повышает их подверженность к окислительной модификации. Клинические исследования выявили значительное превышение уровня КПНГ в ЛНП у больных сахарным диабетом по сравнению со здоровыми лицами. Повышение гликозилирования ЛНП коррелирует с уровнем глюкозы, и содержание КПНГ в ЛНП у пациентов, страдающих диабетом, в 4 раза выше, чем у здоровых лиц [386, 387]. Гликозилирование апо В липопротеинов отмечается преимущественно в положительно заряженных лизиновых участках внутри доменов, связывающихся с рецепторами к ЛНП [387]. Гликирование апо В приводит к значительному снижению их клиренса [1566]. Таким образом, глики- рованные ЛНП плохо распознаются специфическими рецепторами, однако преимущественно узнаются неспецифическими скэвинджер-рецепторами макрофагов [907]. Поскольку гликозилирование ЛНП повышает их поглощение интимой сосудистых клеток и макрофагами с образованием пенистых клеток, узнавание гликированных ЛНП скэ- винджер-рецепторами способствует внутриклеточной аккумуляции эфиров холестерина и прогрессированию атеросклероза. Специфические КПНГ-рецепторы идентифицированы на клеточной поверхности макрофагов, эндотелиальных и гладкомышечных клеток сосудистой стенки [269, 1471].
Вместе с тем гипергликемия может обуславливать и ПОЛ, поскольку известно, что в ЛНП больных сахарным диабетом отмечается значительное повышение содержания гидроперекисей липидов [119], а липопротеины низкой и очень низкой плотности, выделенные из плазмы крови больных диабетом типа II, более подвержены липидной перок- сидации, чем липопротеины здоровых лиц [182, 1652].
Исследованиями Bukala et al. [386] было доказано, что фосфолипиды, составляющие поверхность липопротеиновой глобулы и содержащие в своем составе первичные аминогруппы, значительно быстрее реагируют с глюкозой с образованием КПНГ, нежели полипептиды. Предположив, что внутримолекулярные окислительно-восстановительные реакции, происходящие при формировании КПНГ в фосфолипидах, могут инициировать окисление жирных кислот в составе липопротеинов, авторы показали, что эти процессы идут независимо от добавления металлов с переменной валентностью или экзогенных систем, генерирующих свободные радикалы, доказав тем самым эссенциальную роль процессов гликирования в инициации липидной пероксидации. Было показано при этом, что уровень гликированных липопротеинов в плазме больных диабетом коррелирует с уровнем МДА в ЛНП [386].Процессы ПОЛ и неферментативного гликирования включают аналогичные реакции и промежуточные продукты и ведут к идентичным модификациям лизиновых остатков [385]. Это приводит к мысли, что оба процесса могут усиливать друг друга. АКМ, генерируемые при аутоокислении глюкозы, или ранние и промежуточные продукты гликирования могут инициировать или способствовать пероксидации липидов в липопротеиновых частицах, особенно в условиях гипергликемии [871]. И напротив, повышение уровня окисленных липопротеинов может способствовать связыванию их с глюкозой и образованию КПНГ: так, показано, что неферментативное гликирование может быть усилено гидроперекисями липидов или другими продуктами ПОЛ [755].
Гликированные липопротеины, подвергающиеся окислительной модификации, и КПНГ белков являются химически поврежденными молекулами, следовательно, должны включаться биологические механизмы по их узнаванию и удалению. Как указывалось выше, идентифицированы специфические рецепторы к КПНГ на клеточной поверхности макрофагов, эндотелиальных и гладкомышечных сосудистых клетках [269, 1471]. Связывание КПНГ со специфическими рецепторами сопровождается снижением эндотелиальной барьерной функции и повышением проницаемости базальной мембраны эндотелиоцитов, что приводит к повышению поступления липидов в субэндотелиальное пространство, а окисленные и гликированные ЛНП в свою очередь индуцируют экспрессию молекул адгезии VCAM-1 [1730] с последующей мононуклеарной инфильтрацией интимы [896].
Показано, что связывание КПНГ с рецепторами на клеточной поверхности приводит к развитию окислительного стресса в эндотелиальных и гладкомышечных клетках, характеризующегося активацией пострецепторного сигнала, генерацией внутриклеточных супероксидных анион-радикалов и усилением экспрессии генов [1472, 1798]. Усиление синтеза O2 под действием КПНГ происходит главным образом в результате активации NAD(P)H-оксидаз [1731] (что подтверждается ингибирующим эффектом дифенилениодония), при этом для гладкомышечных клеток показана ведущая роль изоформы Nox1 [1451]. Генерация супероксид-аниона в свою очередь способствует активации транскрипционного фактора NF-κΒ, повышению экспрессии индуцибельной NO-синтазы и развитию нитро- зирующего стресса [1451].Моноциты, инфильтрирующие сосудистую стенку, преобразуются в макрофаги, захватывают окисленные и гликированные ЛНП, превращаясь в пенистые клетки, служащие исходным материалом для формирования атеросклеротической бляшки (см. выше) [363]. Вместе с тем взаимодействие КПНГ с мононуклеарными клетками вызывает продукцию таких иммуномодуляторов, как ИЛ-1, ФНО-α [1702], тромбоцитарный фактор роста [896], инсулиноподобный фактор роста-1 [895], играющих определяющую роль в патогенезе атеросклероза. В частности, показана роль этих факторов в стимуляции гладкомышечной пролиферации сосудистой стенки [1703]. Экспрессия генов этих провоспалительных цитокинов регулируется специфическим ядерным фактором транскрипции NF-κΒ, активация которого происходит при окислительном стрессе [147, 384], в том числе и обусловленном гипергликемией. Была выявлена, в частности, положительная корреляция между активностью NF-κΒ в моноцитах и степенью гликемического контроля (по уровню HbA1c) [768, 769], а у пациентов с диабетической нефропатией — между мононуклеарной NF-кВ-связывающей активностью и выраженностью альбуминурии [769]. Выявлено также значительное подавление активации NF-κΒ, равно как и снижение плазменных маркеров липидной пероксидации, при применении антиоксиданта α-липоевой кислоты у пациентов с сахарным диабетом [768, 769].
NF-κΒ играет ключевую роль в медиации иммунного и воспалительного ответа и апоптоза [384, 1018], и активация этого фактора транскрипции при гипергликемии и вызываемом ею окислительном стрессе подтверждает также воспалительную природу развивающихся ангиопатий [1425].Таким образом, конечные продукты превращений гликозилированных ЛНП способствуют развитию атеросклероза посредством повышения проницаемости эндотелия, усиления адгезии к нему клеток крови, увеличения хемотаксиса моноцитов/мак- рофагов, индукции пролиферации гладкомышечных клеток [312], и гликозилирование ЛНП можно рассматривать как атерогенный фактор, способствующий раннему развитию атеросклероза у больных сахарным диабетом, от осложнений которого погибает до 80 % пациентов.
Среди изоформ супероксиддисмутазы наибольшей способностью взаимодействовать с глюкозой обладают C^Zn-СОД и внеклеточная Э-СОД. Глюкоза связывается с NН2-группами концевых аминокислот или ε-аминогруппами остатков лизина Cu,Zn- СОД, что существенно влияет на ее активность: так, in vitro показано, что культивирование клеток пигментного эпителия сетчатки человека ARPE-19 в условиях гипергликемии сопровождается сдвигом изоэлектрической точки фермента в кислую область за счет выраженного гликозилирования лизиновых остатков, расположенных между а.к.о. 123—143, и снижением ее ферментативной активности в 2,3 раза [1824]. Было обнаружено, что эритроциты человека содержат гликированную и негликированную формы Cu,Zn-SOD, при этом процентное содержание гликированной формы со сниженной ферментативной активностью было выше в эритроцитах у пациентов, страдающих сахарным диабетом, по сравнению с контрольной группой [268]. Возможна также инактивация энзима промежуточными продуктами метаболизма глюкозы (оксальальдегидами): в результате модификации СОД2 метилглиоксалем не только снижается ее ферментативная активность, но и происходит высвобождение ионов меди из активного центра. что может провоцировать образование ОН· [852].
Активность Э-СОД в результате гли- козилирования изменяется незначительно, однако снижается ее сродство к гепарину, в результате чего уменьшается количество фермента, связанного с гепаринсульфатом гликокаликса эндотелия, и ослабляется антиоксидантная защита эндотелиоцитов [65], при этом более легкая «десквамация» гликированой Э-СОД не сопровождается увеличением концентрации фермента в кровотоке, поскольку ускоряется деградации модифицированного фермента макрофагами печени, и в результате внутривенной инфузии гепарина активность Э-СОД в циркулирующей крови больных диабетом возрастает в 1,88 раза (в группе сравнения — в 1,5 раза), оставаясь существенно ниже, чем у здоровых людей (13,5 ± 3,3 ЕД/мл и 20,3 ± 1,7 еД/мл соответственно) [470].Присоединение остатка глюкозы к молекуле трансферрина снижает его способность связывать ионы железа, одновременно гликозилирование связанных с ионами молекул трансферрина приводит к тому, что они становятся источниками образования О- и ОН· [628]. Антиоксиданты (витамин Е, витамин С, пробукол) снижали накопление продуктов ПОЛ в сыворотке и тканях, а также защищали белки от повреждения, вызванного гликозилированием, как у больных с диабетом, так и у экспериментальных животных [154, 868, 1427].
У здоровых людей гликировано около 5 % гемоглобина, при высокой гипергликемии это значение может возрастать до 15-20 %, превышая нормальные значения в 2-3 раза. В лабораторной практике зону концентраций гликогемоглобина 6-7 % принято считать пограничной, при концентрациях в этих пределах диагностируют «подозрение на диабет». Превышение десятипроцентного порога считают однозначным признаком декомпенсированного диабета. Гликозилированный гемоглобин теряет многие свои функциональные свойства, в том числе способность транспортировать кислород, что может приводить к развитию тканевой гипоксии, одной из основных составляющих патогенеза сахарного диабета (наряду с энергетическим голодом, клетки испытывают недостаток кислорода). Тканевая гипоксия приводит к повышению в крови уровня лактата (молочная кислота), который образуется в процессе анаэробного расщепления глюкозы в клетках. Высокий уровень лактата наблюдается примерно у половины больных сахарным диабетом, при этом наибольшие концентрации лактата в сыворотке выявляются при интенсивных физических нагрузках. На культурах гладкомышечных клеток сосудов крыс было показано, что гликозилированный гемоглобин может индуцировать клеточную гипертрофию. Культивирование в течение 48 ч гладкомышечных клеток с гликозилированным гемоглобином человека (гликозилировано 14 %) в концентрациях выше 10 нМ сопровождалось увеличением клеточной поверхности, которое подавлялось СОД (200 ЕД/мл), дефероксамином (100 нМ — 100 мкМ), диметилтиомочевиной (1 мМ) и частично каталазой (200 ЕД/мл) и маннитолом (1 мМ) [1301]. Аскорбиновая кислота и ряд лекарственных препаратов (гликлазид, нифедипин и др.), обладающие антиоксидантными действиями, также ингибировали индуцированную гликозилиро- ванным гемоглобином гипертрофию гладкомышечных клеток [1302].
Еще по теме Конечные продукты неферментативного гликирования белков и липидов:
- Наличие научно обоснованных требований к результатам и «конечному продукту» функционирования каждого
- Деструкция белков
- Изменения белков
- Неферментативные природные и синтетические антиоксиданты
- Денатурация белков
- Классификация, состав и структура липидов мембран
- Физико-химические и динамические свойства и функции липидов мембран
- ВЫДЕЛЕНИЕ И ОЧИСТКА МЕМБРАННЫХ БЕЛКОВ
- Неравновесные состояния белков и их релаксация
- Метаболизм углеводов и липидов
- 1.2.1. Классификация, структура и функции мембранных белков