Неравновесные состояния белков и их релаксация
В литературе имеется много работ, посвященных конформацион- но неравновесным состояниям белков (ферментов) после локального возмущения. Помимо простой регистрации, была исследована кинетика релаксации, а также их физические и химические свойства в промежуточных состояниях во время конформационной релаксации.
Большинство работ, выполненных в разных лабораториях, использовали белки с ионами металлов в их активных центрах. Это облегчает использование техники электронного парамагнитного резонанса (ЭПР), оптического поглощения и т. д.Есть два главных метода получения металлсодержащих белков в неравновесных состояниях.
Для получения и сохранения неравновесных форм металлсодержащих белков и исследования их спектральных и магнитных свойств была разработана специальная техника низкотемпературного восстановления [38]. В этом методе содержащая металл простетическая группа1^ восстанавливается в замороженном водном растворе при температуре жидкого азота с помощью радиолиза. При этом образуются термолизованные2) электроны. Исходные препараты представляют собой белки, активные центры которых могут существовать как в окисленной, так и в восстановленной формах.
После электронного восстановления активного центра возникают кинетически стабилизированное неравновесное состояние. Атом металла в активном центре восстановлен, но его непосредственное окружение изменено настолько, насколько это позволяет замороженная в матрице и поэтому не изменившаяся глобула. Непосредственное окружение активного центра претерпевает колебательную релаксацию, но пространственная структура глобулы остается той же, какой она была в равновесном окисленном белке. Однако новое состояние активного центра и его ближайшего окружения должно в условиях равновесия соответствовать конформации всей белковой глобулы. Напряжение между измененным активным центром и остальными частями макромолекулы приводит к изменению спектральных и магнитных характеристик активного центра.
Так возникает конформационно неравновесное состояние белка: ион металла восстановлен, но структура большей части глобулы соответствует окисленному состоянию иона металла. Методы получения, фиксирования и исследования конформационно неравновесных состояний могут с успехом использоваться не только для отдельных белков, но и для внутриклеточных органелл, клеток и целых тканей.С повышением температуры можно наблюдать релаксацию не ре- лаксировавшей части глобулы к новому равновесному состоянию.
«Простетическая» — небелковая часть фермента. Обычно играет роль активного центра.
«Термолнзованные* — находящиеся в термодинамическом равновесии с термостатом.
Температура релаксации определяется природой белка и свойствами матрицы.
Конформационно неравновесные состояния белков (и не только металлсодержащих) могут быть получены в растворе при комнатной температуре с помощью следующих методов:
i. Восстановление иона металла микросекундными (или более короткими) импульсами быстрых электронов или ароматическими анион-радикалами, образуемыми из соответствующих соединений с помощью вспышек света короткой длительности.
ii. Флаш-фотолиз гем-содержащих белковых комплексов с О2, CN-,
N0.
iii. Скачкообразное добавление субстрата, ингибитора или другого специфического лиганда к равновесному белку, а также скачкообразное изменение pH или ионной силы.
Мы выберем в качестве примера «почетный фермент» —- гемоглобин. Строго говоря, гемоглобин не является ферментом, его функция
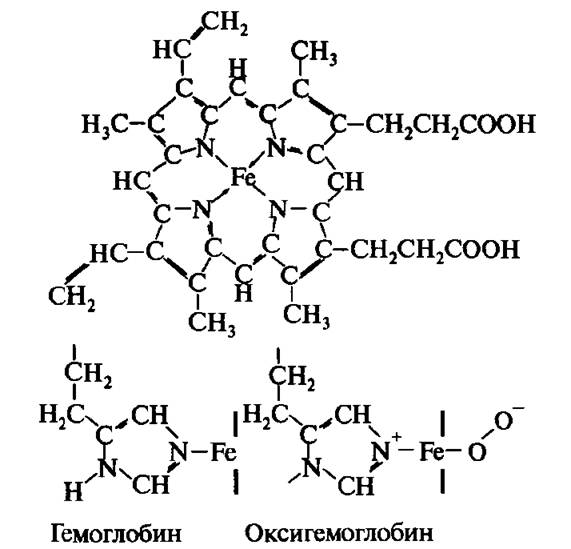
Рис. 4.8. Упрощенная схема активного центра гемоглобина. Структура гема и присоединение кислорода к железу гема
заключается в переносе О2 от легких к тканям и СО2 от тканей к легким. После локального возмущения, однако, он ведет себя точно так же, как и ферменты.
Рис. 4.8 представляет упрощенную схему молекулярной структуры гемоглобина.
Молекулы гемоглобина состоят из 4-х субъединиц: двух a и двух р и, соответственно, содержат 4 полипептидные цепочки двух типов. Каждая а-цепочка содержит 141, а /3-цепочка —
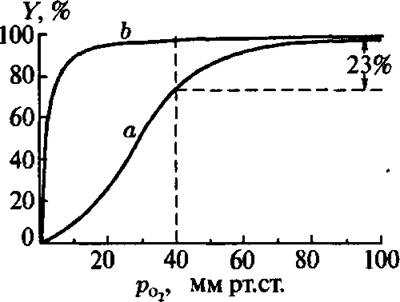
Рис. 4.9. Кривые оксигенации гемоглобина (а) и миоглобина (Ь)
146 аминокислотных остатков. Вся молекула гемоглобина содержат, таким образом, 574 аминокислот.
Строго говоря, эти детали структуры относятся не к гемоглобину, а к его белковому компоненту — глобину. Каждая субъединица глобина содержит одну небелковую (про- стетическую) группу — гем: комплекс порфирина с Fe2+. Структура гема показана в верхней части рис. 4.9. Атом железа может образовать шесть координационных связей. Четыре связи направлены к атомам азота порфиринового кольца, а оставшиеся четыре связи — перпендикулярны к плоскости кольца по обе ее стороны. Гемы расположены вблизи поверхности белковой глобулы в специальных «карманах», образованных складками полипептидных цепочек глобина. Нормально функционирующий гемоглобин существует в одной из трех форм: феррогемоглобин (называемый также дезоксигемоглобином или просто гемоглобином), оксигемоглобин и ферригемоглобин (метгемоглобин). Ион железа в феррогемоглобине находится в восстановленной форме Fe2+, одна из двух связей, перпендикулярных к плоскости кольца, соединяет железо с атомом азота гистидинового остатка, другая связь свободна (нижняя часть рис. 4.9). Взаимодействие молекулярного кислорода со свободным гемом приводит к окислению иона железа
(Fe2+ ------- >Fe3+). В случае феррогемоглобина глобин предохраняет
железо гема от окисления.
Мышечные ткани содержат миоглобин, который, грубо говоря, является четвертушкой гемоглобина. Миоглобин имеет только одну субъединицу, структура которой идентична структуре ft субъединицы гемоглобина. Миоглобин связывает кислород значительно сильнее, чем гемоглобин, и служит кислородным депо при недостатке кислорода.
Обратимое присоединение кислорода (оксигенация), которое позволяет гемоглобину выполнять его основную функцию, обусловлено возможностью образования сильных пятой и шестой координационных связей и способностью переносить электрон к кислороду от гистидина, а не от железа, т.е. без окисления Fe2+ (см. рис. 4.8).
Как уже было сказано выше, молекула гемоглобина содержит четыре субъединицы и, следовательно, четыре гема, каждый из которых может присоединить одну молекулу кислорода. Поэтому реакцию оксигенации можно разделить на четыре стадии:
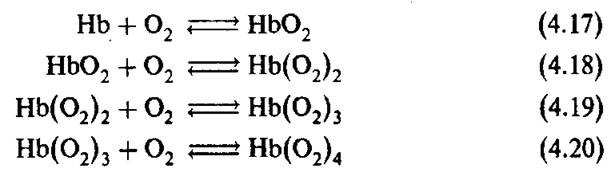
На рис. 4.9 представлена типичная S-образная кривая оксигенации гемоглобина (кривая а) и гиперболическая кривая оксигенации миоглобина (кривая Ь). Последняя соответствует более прочному связыванию кислорода активным центром миоглобина.
В отличие от миоглобина, гемоглобин на начальных стадиях оксигенации обладает низким сродством к кислороду, и равновесие в реакции (4.17) сдвинуто влево. Затем кривая становится круче и при высоких значениях рО2 достигает насыщения. Это типичный эффект кооперативности.
Активность многих ферментов скачкообразно изменяется от одного значения к другому под действием некоторых низкомолекулярных агентов, которые не принимают непосредственного участия в каталитическом акте [39].
Это явление называется аллостерическим эффектом. Такие ферменты могут существовать в разных конформациях с различными активностями. Эти конформации находятся в динамическом равновесии. Присоединение низкомолекулярного агента смещает равновесие
и меняет поэтому активность фермента. К этому же классу белков принадлежит и гемоглобин.
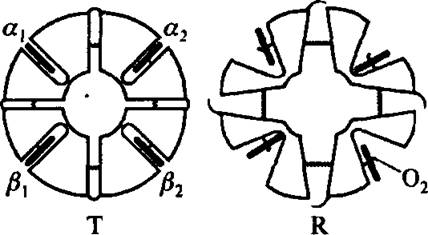
Рис. 4.10. Четвертичная структура молекулы гемоглобина
На рис, 4.10 представлена схема четвертичной структуры двух конформационных состояний гемоглобина.
Показаны только изменения в относительном расположении четырехсубъединиц. В форме Т (от английского “tense”) гемы расположены в узких карманах глобулы и присоединение кислорода затруднено. В форме R (от английского “rapid”) атомы железа гема открыты и присоединение кислорода облегчается.
Конформационный переход Т ------------ ► R включает не только измене
ние четвертичной структуры белковой глобулы. Релаксация затрагивает все части четырех субъединиц. Чтобы понять последовательность событий в ходе релаксационного процесса, рассмотрим основные физические характеристики трех главных форм гемоглобина. Они приведены на рис. 4.11.
В равновесном гемоглобине ион железа (Fe2+) лежит вне пор- фиринового кольца (примерно на 1 А). Он имеет четыре электрона и магнитный момент, равный 5,5 Боровских магнетонов. Оптический спектр поглощения имеет широкую полосу с Лмах = 5,56 нм. В равновесном оксигемоглобине ион железа (Fe2+) находится точно в плоскости порфиринового кольца, все электроны спарены (оксигемоглобин диамагнитен). В спектре оптического поглощения видны две характеристические полосы при 542 и 576 нм. В ферригемоглобине (метгемоглобин) при нейтральных значениях pH молекула кислорода замещается молекулой воды, не связанной химически с ионом железа (Fe3+). Ион железа лежит значительно ближе к порфириновому кольцу, чем в феррогемоглобине (почти в плоскости), имеет пять неспаренных электронов и магнитный момент равный 5,91 Боровских магнетона. Спектр поглощения в видимой области не имеет выраженных характеристических полос.
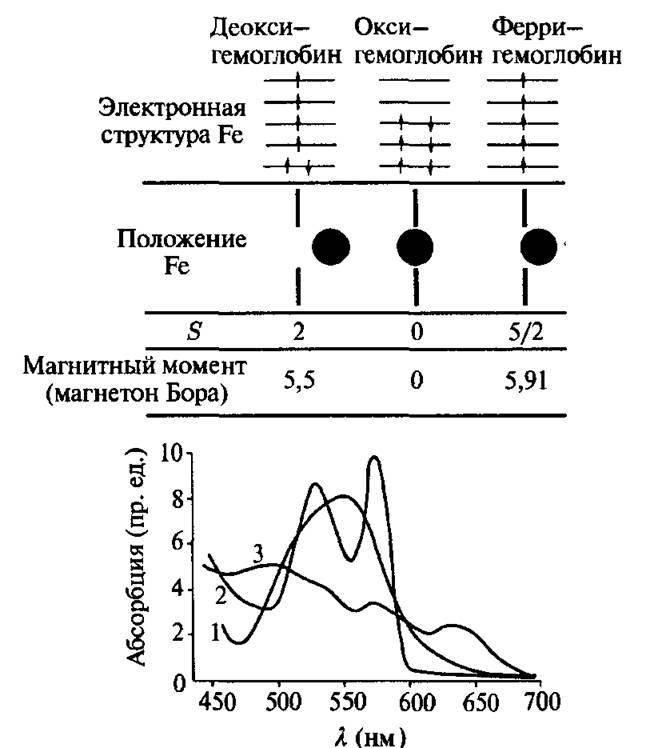
Рис. 4.11. Физические свойства основных производных гемоглобина
Структурные изменения в активном центре (вблизи гема) приводят также к значительным изменениям пространственной структуры всего белка. После присоединения кислорода (Т ► R-переход), некото
рые аминокислотные остатки сдвигаются на ~ 7 А.
Эти структурные изменения инициируются присоединением одной молекулы кислорода, а затем распространяются на всю белковую глобулу. Поэтому в равновесной смеси присутствуют только Т и R формы.Рассмотрим последовательность во времени таких структурных изменений, используя в качестве примера быстрое электронное восстановление ферригемоглобина.
Прежде всего подведем итог и суммируем основные структурные характеристики равновесных форм главных производных гемоглобина.
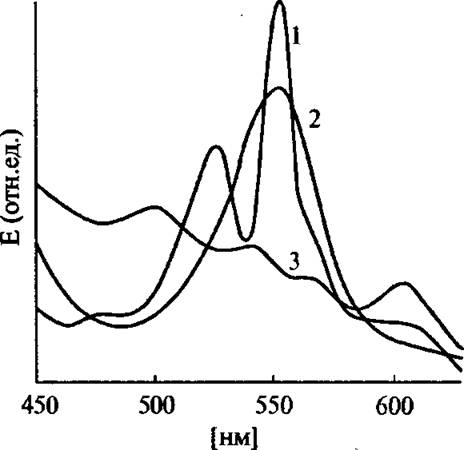
Рис. 4.12. Спектры поглощения неравновесного феррогемоглобина (1), равновесного феррогемоглобина (2) и равновесного ферригемоглобина (3) в замороженных растворах при 77 К
В феррогемоглобине пентакоординированный ион Fe2+ находится в высоко-спиновом состоянии (S = 2) и значительно смещен от плоскости порфиринового кольца. Присоединение О2 (как и СО или N0) сопровождается переходом железа гема в низко-спиновое состояние (S = 0) и смещением атома железа в плоскость порфиринового кольца. В то же время структура активных центров в равновесном высоко-спиновом ферригемоглобине такова же, как и в низко-спиновом феррогемоглобине: ион железа находится почти в плоскости порфиринового кольца. Это же справедливо и для соответствующих производных миоглобина. Подробное теоретическое обсуждение этих структурных характеристик можно найти в [40].
После восстановления ферригемоглобина в замороженной матрице быстрыми электронами более 80 % белка находится в неравновесной восстановленной низко-спи
новой форме, характеризу- ющейся спектром поглощения типа «оксигемоглобина»
(рис. 4.12). Повышение температуры приводит к релаксации неравновесного феррогемоглобина в равновесное состояние со спектром, показанным на рис. 4.12 (кривая 2). Аналогичные эффекты наблюдались для комплексов ферригемоглобина с различными лигандами: фтор, азид, цианид, имидазол или ОН- (pH 8,5). Восстановление этих комплексов в замо- ,[х>женных растворах приводит к образованию со 100%
выходом неравновесных восстановленных форм белка со спектрами поглощения и магнитного кругового дихроизма соответствующих низко-спиновых равновесных форм производных феррогемоглобина.
Анализ этих данных говорит о том, что образование низко-спиновых неравновесных форм феррогемоглобина и его комплексов, после низкотемпературного восстановления ферригемоглобина и его комплексов, обусловлено сохранением аксиальных лигандов в шестом координационном положении иона Fe2+. Это положение, по всей видимости, занято атомом азота в имидозольном остатке дистального гистидина. Диссоциация лиганда блокирована стерическими препятствиями белковых групп вокруг гема.
Те же результаты были получены с ферримиоглобином и его комплексами.
Образование неравновесных форм многих металлсодержащих белков в замороженных растворах быстрыми электронами было зарегистрировано при низкотемпературном восстановлении с использованием различных физических методов (оптическое поглощение, ЭПР, магнитный круговой дихроизм). Все ссылки можно найти в монографиях [1,4,31].
Как уже было отмечено, конформационно неравновесные состояния всех исследованных белков с повышением температуры релак- сируют к соответствующим равновесным состояниям еще до размораживания матрицы. В некоторых случаях удалось установить последовательность структурных перестроек. Было найдено, например, что перед удалением аксиального лиганда из координационной сферы атома железа его ориентация меняется и атом железа удаляется от порфиринового кольца. Дальнейшая релаксация проходит через несколько промежуточных состояний.
Данные об образовании конформационно неравновесных состояний железосерных белков, играющих важную роль во внутриклеточной трансформации энергии, будут рассмотрены позже. В конце этого параграфа будут описаны различные методы получения неравновесных форм белков в растворах при комнатной температуре. До настоящего времени этой техникой было исследовано несколько сотен белков, зарегистрирована кинетика их релаксации и установлены их физические и химические свойства. Здесь будут даны только принципиальные выводы из результатов этих исследований. Ссылки на соответствующие публикации можно найти в книгах [1,31,37,41].
Анализ приводит к следующим важным выводам:
1. Во всех случаях быстрого локального возмущения активного центра белка (фермента) возникает неравновесное состояние, которое релаксирует путем конформационных изменений, охватывающих всю белковую глобулу. В ходе этих изменений молекула белка проходит через множество промежуточных, неравновесных состояний, переходы между которыми могут занимать от микросекунд до сотен миллисекунд. Специфическая химическая активность молекулы белка (например, ферментативная активность), как правило, в этих промежуточных неравновесных состояниях выше, чем в исходном и конечном равновесных состояниях.
2. Акт превращения субстрата в продукт совпадает с одной из стадий конформационной релаксации белка.
Несколько примеров. Для многих белков были измерены при комнатной температуре характерные времена различных стадий релаксации после скачкообразного восстановления иона железа в активном центре [41]. Для цитохрома С (один из главных компонентов дыхательных электрон-транспортных цепей в митохондриях аэробных организмов), при pH 10,6 были зарегистрированы три релаксационные стадии с характерными временами, около 50 мкс, 0,5 мс и 0,3 с. Для гемоглобина при pH 7,4 масштабы времени обнаруженных релаксационных стадий были: 70 мкс, 0,3 мс, 0,6 мс и 50 мс. Для железосерного белка — фер- редоксина были зарегистрированы две четкие стадии с характерными временами около 100 мкс, 0,2 с.
Достаточно большие времена жизни металлсодержащих белков в конформационно неравновесных состояниях делают возможным оценить их химическую реакционную способность. Измерили константы скорости некоторых специфических реакций этих белков сразу после образования их неравновесных состояний и в течение релаксации. Были подробно исследованы следующие реакции: окисление восстановленных железосодержащих белков феррицианидом калия или некоторыми металлсодержащими белками в окисленной форме.
Главным результатом этих кинетических исследований было то, что константы скорости реакций белков в неравновесных состояниях могут отличаться существенно (вплоть до 103 раз) от соответствующих констант скорости тех же белков в равновесных состояниях. Реакционная способность обычно повышена (но не всегда). После релаксации константы скорости возвращаются к значениям, характерным для равновесных белков.
Скорости реакций неравновесных белков часто имеют необычную зависимость от температур. Начальная конфигурация и скорость конформационной релаксации белков, как правило, более чувствительны к изменениям температуры, чем скорость элементарного акта исследуемого химического превращения. Поэтому, например, с понижением температуры конформационная релаксация замедляется и большая часть реакции успевает закончиться на первых стадиях релаксации, когда реакционную способность белка максимальна. Измеряемая скорость реакции в этом случае возрастает в кажущемся противоречии с законом Аррениуса.
Описанные выше исследования белков в конформационно неравновесных состояниях требуют сложной экспериментальной техники: низкие температуры для замедления релаксации, импульсные методы для одновременной реализации локальных химических изменений в значительной части молекул белка в образце и т.д. Совершенно ясно, однако, что аналогичные события могут иметь место и с отдельными молекулами белка во время их функционирования при физиологических температурах, без искусственно навязанной синхронизации. Полученные до сих пор данные доказывают, что многие реакции белков действительно протекают в две главных стадии: быстрые локальные изменения с последующей медленной релаксацией, в ходе которой белковые молекулы проходят через серии существенно неравновесных состояний и реализуют (для ферментативных процессов) трансформацию субстрата в продукт.
Релаксация неравновесного состояния, специально приготовленного с помощью направленного, локального воздействия, может быть с одинаковой степенью обоснованности описана либо как последовательностью элементарных актов, каждый из которых протекает обратимо и переводит молекулу в новое конформационное состояние (см. рис. 4.6 и 4.7), либо как непрерывное движение вдоль выделенной механической степени свободы. В случае достаточно простых молекул предпочтителен первый подход. Во время релаксации сложных
белковых молекул элементарные акты (повороты вокруг простых ковалентных связей, изменения некоторых валентных углов, разрыв и образование водородных связей и т. д.) должны быть реализованы в строго определенной последовательности с помощью правильных и ошибочных элементар ных актов. Если число возможных элементарных актов достаточно велико, то описание релаксационного процесса становится неудобным, практически невозможным и поэтому бессмысленным, как, например, микроскопическое описание любого макроскопического движения.
Исследование превращения субстрата в продукт и конформационной релаксации во время одного оборота ферментативной реакции [42]. Исследовали прямую и обратную реакции, катализируемые растворимым ферментом малатдегидрогеназой (МДГ) с коферментом NAD:
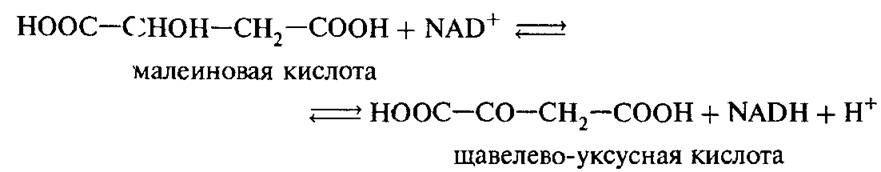
Конформационные изменения МД Г изучали, измеряя (пикосекунд ,ная техника) среднее время жизни собственной флуоресценции триптофана (ту), которая чувствительна к его непосредственному окружению. Значение Ту измеряла каждые 0,5 мс в течение 1,5 оборота белка (~ 10 мс для прямой реакции). Химическую трансформацию субстрата детектировали по изменениям редокс состояния кофермента, измеряя оптическую плотность образца при 340 нм (максимум поглощения NADH) каждые 0,5 мс, начиная с 0,5 мс после смешивания. Присоединение L-малата или оксалацетата не меняет значения Ту МД Г, но образование комплексов с коферментами NAD+ или NADH понижает Ту при 20° С от примерно 3,6 нс до ~ 3,1 нс в течение ~ 4 мс после смешивания. На рис. 4.13 а представлены значения Ту и кинетика восстановления NAD+ при 20° С в течение 14 мс после смешивания. Если реакция была инициирована добавлением L-малата к смеси МД Г и NAD+, Ту немедленно начинает уменьшаться, достигая минимального значения (~ 2,5 нс) при ~ 6^0 мс после смешивания. В конце первого цикла (через ~ 10,5 мс) Ту возвращается к своему
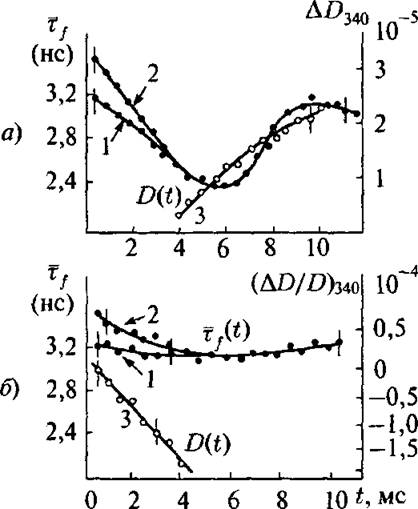
Рис. 4.13. а) Кинетика изменений оптической плотности NADH, DM0 (кривая 3) и изменений ту после инициирования прямой реакции. Кривая 1: реакция инициирована добавлением субстрата к смеси фермент + кофермент. Кривая 2: реакция инициирована добавлением смеси кофермент + субстрат к ферменту. 6} Кинетика изменений D340 и ту после инициирова ния обратной реакции. Обозначение те же, что и на рис. 4.13 а
первоначальному значению, характерному для МД Г—NAD+, а затем снова уменьшается, хотя и медленнее., чем в начале первого цикла. Если реакция была инициирована добавлением NAD+ к смеси МДГ с L-малатом или добавлением смеси NAD+ с L-малатом к МД Г, то кривая Ту (0 в течение первых 4 мс проходит выше соответствую щей кривой для первой схемы опыта, после чего обе кривые совпадают'.
При 5° С для обеих схем и инициаций Ту уменьшается в течение 14 мс после смешивания и не достигает своего минимального значения до конца регистрации.
Совершенно другие результаты были получены для обратной ре акции (восстановление оксалацетата и окисления NAD+). Эти результаты показаны на рис. 4,13 Реакция практически не сопровождается конформационной релаксацией, регистрируемой по уменьшению Ту, а окисление NAD+ начинается сразу после смешивания и продолжается в течении нескольких оборотов фермента (время оборота фермента для обратной реакции примерно 3 мс).
Все эти результаты можно рассматривать как доказательства того, что МДГ катализирует прямую и обратную реакции, находясь в различных конформационных состояниях. Пути этих реакций не совпадают.
Эти данные могут также прояснить механизм «гистерезиса» или «памяти» ферментативной активности. Существуют две возможности гистерезисного поведения таких белков.
1. Постепенное возрастание активности каждой молекулы фермента с каждым новым оборотом.
2. Возрастание числа активных молекул белка в популяции первоначально неактивных молекул фермента.
Из выше представленных данных следует, что в первом цикле оборота фермента участвует лишь незначительная часть молекул МДГ (~ 0,1 %), но все эти молекулы обладают максимальной активностью с самого начала. Это типичное поведение гистерезисных ферментов, которые могут существовать в двух дискретных состояниях — активных и неактивных.
В ходе реакции число активных молекул возрастает. Каждая активная молекула фермента, принимая участие в акте превращения субстрата в продукт, выводится из популяции потенциально активных, но не реагировавших молекул. Поэтому равновесие между двумя популяциями смещается и число активных молекул фермента возрастает.
4.7.