§ 11. Библиография и комментарии
§ 1. Существует огромное число работ, в которых и по результатам наблюдений за природными системами, и на искусственно конструируемых лабораторных системах изучалась динамика численностей двух видов, связанных взаимоотношениями типа хищник — жертва.
Наиболее известные работы — это наблюдения за колебаниями численностей рысей и зайцев в Канаде (в течение почти ста лет) и лабораторные эксперименты 30-х годов Г. Гаузе с двумя популяциями простейших. Из сравнительно недавних работ можно указать на очень большие по объему эксперименты Утиды:U t і d a S. Cyclic fluctuations of population density intrinsic to the host-parasite system. — Ecology, 1957, 38, № 3, p. 442— 449.
Им было поставлено несколько десятков экспериментов, но тем не менее не удалось обнаружить каких-либо общих закономерностей в поведении системы паразит — хозяин, кроме, быть может, наличия колебаний численностей и сдвига по фазе. Некоторые из экспериментальных систем быстро гибли, другие существовали довольно долго, причем колебания сглаживались.
Сравнительно недавно появился цикл работ, в которых большое внимание было уделено изучению различных механизмов регулирования численности жертвы в этих системах (в том числе механизмов, связанных с пространственным распределением видов и с их поведенческими реакциями):
White Е. G., Н u f f a k е г С. В. Regulatory processes and population cyclity in laboratory populations of Anagasta kuhniella. II. Parasitism, predation, competition and protective cover. —Res. Pop. Ecol., 1969, II, № 2, p. 150—155; Matsumoto В. M., Huf faker С. B. Regulatory processes and population cyclity in laboratory populations of Anagasta kuhniella. V. Host finding and parasitization in a
«small» universe by an entomophagous parasite, Venturia canescens. —Res. Pop. Ecol., 1974, 15, p 193—212; VI. Host finding and parasitization in a «large» universe by an entomophagous parasite, Venturia canescens.
—Res. Pop. Ecol., 1974, 15, p. 213—234.В работе
Pimentel D. Population regulation and genetic feedback. — Science, 1968, 159, № 3822, p. 1432—1437
было предложено новое объяснение механизма возникновения устойчивости в системе паразит — хозяин за счет генетической коэволюции этих двух популяций. Но модели такого типа выходят за рамки нашего изложения.
§ 2. Понятие функционального отклика (трофической функции) было впервые введено Холлингом в его известных экспериментальных работах по изучению процесса хищничества:
Н о 1 1 і n g С. S. The functional response of predator to prey density and its role in mimicry and population regulation.— Mem. Entomol. Soc. Canada, 1965, 45, p. 1—60.
В работе Холдинга S-образные трофические функции получались при весьма жестких требованиях на рацион хищника, однако Рояма в работе:
R оу a ma Т. A comparative study of models of predation and parasitism. —Res. Pop. Ecol., 1971, Supp. 1, p. 1—91 показал, что эти ограничения можно ослабить. И, наконец, в работе
Hassel М. Р., Lawton J. Н., Beddington J.R.
Sigmoid functional responses by invertebrate predators and parasitoids. —J. Anim. Ecol., 1977, 46, № 1, p. 249—262
были проведены многочисленные экспериментальные данные, из которых следует, что как раз S-образная трофическая функция является наиболее распространенной в реальных природных системах, причем не только у позвоночных, но и у беспозвоночных хищников и паразитов.
Существуют, однако, эксперименты, в которых получены трофические функции и весьма экзотического вида; например, неограниченно возрастающая функция, несколько раз меняющая знак своей кривизны (см.
Haynes D. L., Sisojevic Р. Predator behavior of Philodromus rufus Walckenaer. —Can. Entomol., 1966, 98, № 1, p. 113—133).
§ 3. Здесь мы воспользовались известной теоремой Барбашина — Красовского об устойчивости в целом или глобальной устойчивости:
Барбашин Е. А. Введение в теорию устойчивости. —
М.: Наука, 1967, Гл. 1.
О понятии «грубости» динамических систем см,, например,
Баутин Н.
Н., Леонтович Е. А. Методы и приемы качественного исследования динамических систем на плоскости. — М.: Наука, 1976.§ 4. Впервые обобщение вольтерровской модели было предложено А. Н. Колмогоровым в статье:
Kolmogorov A. Sulla teoria di Volterra della lotta per l’esistenze. — G. Inst. Ital. Attuari, 1936, 7, № 1, p. 74— 80.
Переработанный и расширенный вариант этой работы:
Колмогоров А. Н. Качественное изучение математических моделей популяций. —М.: Наука, 1972, с. 100—106. — (Проблемы кибернетики; Вып. 25).
Модели аналогичного типа рассматривались в статьях:
Rosenzweig М. L., MacArthur R. Н. Grafical representation and stability conditions of predator-prey interactions. — Amer. Natur., 1963, 97, № 893, p. 209—223; Rescigno A. The struggle for life: 1. Two species. — Bull. Math. Biophys., 1967, 29, № 2, p. 377—388.
Естественно, что модель Колмогорова, обладая богатым набором различных фазовых траекторий, могла качественно описать практически любую реальную ситуацию, но ее излишняя общность затрудняла как количественную идентификацию модели, так и интерпретацию полученных результатов. Поэтому появилось достаточно много моделей, занимавших промежуточное положение между вольтерровской и колмогоровской моделями. В известной мере к ним можно отнести и уже цитировавшуюся модель Розенцвейга и Мак-Артура, которая в наших обозначениях имеет вид
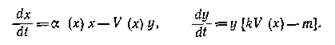
Она была подробно исследована Фридманом:
Freedman Н. I. Graphical stability, enrichment and pest control by a natural enemy. —Math. Biosci. 1976, 31, № 3/4, p. 207—225,
который получил условия устойчивости нетривиального равновесия. Но наиболее интересный его результат — это достаточные условия существования предельного цикла.
Различные типы поведения фазовых траекторий для трофической функции I и при наличии самолимитирования либо популяции жертвы, либо хищника, либо обеих вместе были подробно исследованы в работе:
Б а з ы к и н А.
Д. Система Вольтерра и уравнение Миха- элиса—Ментена. — В кн.: Вопросы математической генетики. Новосиб ирск, СО АН СССР, 1974, с. 103—143.Самолимитирование описывалось членами вида — ул-х2 и — УуУ2-
§ 5. Модель такого типа (а(х) = const) была впервые предложена в работе
Gause G. F., S m а г a g d о v а N. Р., W 1 11 А. А. Further studies of interaction between predator and prey. — J. Anim. Ecol., 1936, 5, № 1, p. 1—18
для интерпретации экспериментов Гаузе с лабораторными системами хищник — жертва.
§ в. В уже цитировавшейся статье Фридмана, а также в работах
Freedman Н. I., Waltman Р. Perturbation of two- dimensional predator-prey equations. —SIAM J. Appl. Math., 1975, 28, № 1, p. 1—10;
Freedman H. I., Waltman P. Perturbation of twodimensional predator-prey equations with an unperturbed critical point. — SIAM J. Appl. Math., 1975, 29, № 4, p. 719—733 для систем, близких к вольтеооовским. т. е. для систем вида
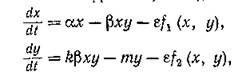
были получены достаточные условия существования устойчивого предельного цикла. Вообще говоря, они сразу следуют из бифуркационной теоремы Хопфа (см., например,
Ч е з а р е Л. Асимптотическое поведение и устойчивость решений обыкновенных дифференциальных уравнений. — М.: Мир, 1964, Гл. III, § 8.6),
но они получаются настолько громоздкими, что даже их проверка представляет собой весьма сложную задачу. Кроме того, здесь трудно дать содержательную интерпретацию малому параметру в. Например, если трофическая функция V(x) ограничена, то при достаточно больших х V(x) будет значительно отличаться от линейной функции VB = —
трофической функции вольтерровской модели. В то же время, если перейти к переменным (£, Т)), то понятие близости становится достаточно очевидным и появляется возможность наглядной интерпретации малого параметра, а теорема Понтрягина позволяет получить условия существования устойчивого предельного цикла в простой и биологически содержательно интерпретируемой форме.
Цитируемая здесь теорема Понтрягина впервые изложена в статье:
Понтрягин Л. С. О динамических системах, близких к гамильтоновым. —ЖЭТФ, 1934, 4, Вып. 8.
§ 7. Изложение метода Крылова — Боголюбова см. например, Моисеев Н. Н. Асимптотические методы в нелинейной механике. — М.: Наука, 1969.
Заметим, что аналогичное решение можно было получить и прямо из (6.8) или (6.9), взяв в качестве (или Sc) эллипс и разложив подынтегральную функцию в ряд Тейлора до соответствующих степеней. Поскольку h* (или С*) выражается через длины полуосей этого эллипса, то уравнение ф(/г*) = 0 (ф(С*) = 0) есть не что иное, как уравнение для нахождения стационарных амплитуд.
Конкретное представление трофической функции типа III в виде
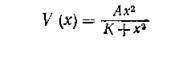
было предложено в работе:
Н a s s е 1 М. Р., V а г 1 е у G. С. New inductive population model for insect parasites and its bearing on biological control. — Nature, 1969, 223, № 5211, p. 1133—1137.
§ 8. Реально наблюдавшийся эффект «ускользания» в системе хищник — жертва из двух популяций насекомых описан в работе:
Takanashi F. Reproduction curve with two equilibrium points: a consideration on the fluctuation of insect population. — Res. Pop. Ecol., 1964, 6, № 1, p. 28—36.
§ 9. О применении метода Крылова — Боголюбова для нахождения параметрического резонанса в нелинейных колебательных системах со случайными параметрами см.
Митропольский Ю. А. Метод усреднения в нелинейной механике. — Киев: Наукова думка, 1971.
Техника, используемая в этом параграфе, изложена в книге:
К л я ц к и н В. И. Статистическое описание динамических систем с флуктуирующими параметрами. —М.: Наука, 1975,
в 6 главе которой подробно исследована (причем различными методами) система (9.3). По-видимому, эта техника может быть использована и для изучения поведения моделей более сложных сообществ при случайных флюктуациях их параметров типа «белого шума».
Еще по теме § 11. Библиография и комментарии:
- § 7. Библиография и комментарии
- § 9. Библиография и комментарии
- § 8. Библиография и комментарии
- § 8. Библиография и комментарии
- § 9. Библиография и комментарии
- § 9. Библиография и комментарии
- § 9. Библиография и комментарии
- § 15. Библиография и комментарии
- § 4. Отраслевая библиография
- Библиография
- 2. 1. Библиографии