УФ-индуцированные изменения функциональных свойств мем-брапосвязанной Na+, КР-АТФазы в присутствии фосфолипазы D
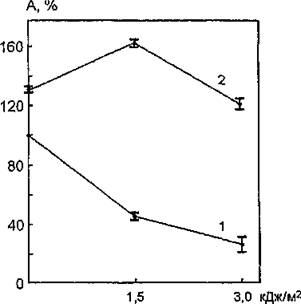
На рис. 48 представлены данные по исследованию фоточувствительности мембраносвязанной Na+, К+-АТФазы при воздействии УФ-излучения (240—390 нм) в дозах 1,5 и 3,0 кДж/мй на интактные мембраны эритроцитов человека.
Из анализа рисунка следует, что при облучении мембран происходит статистически достоверное снижение уровня каталитической активности исследуемого фермента.Рис. 48. УФ-индуцированные изменения функциональной активности Na+, К+-АТФазы эритроцитарных мембран, модифицированных фосфолипазой D: 1 — УФ-облучение интактных мембран; 2 — УФ-облучение модифицированных фосфолипазой мембран
В состав активного центра Na+, К+-АТФазы входят высокореакционноспособные остатки тирозина и цистина, поглощение УФ-света которыми обусловливает процесс фотомодификации фермента.
Воздействие длинноволнового УФ-света в интервале длин волн 320—390 нм (Xmnx - 365 нм) в течение 30 мин приводит к полному ингибированию ферментативной активности Na+, К+-АТФазы. Хромофорами УФ-света в этом диапазоне длин волн являются различные (восстановленные) пиридиннуклеотиды, флавины, железопор- фирины. Таким образом, инактивация мембраносвязанного фермента в данном случае может быть обусловлена фотохимическими превращениями вышеназванных хромофоров. Не исключена вероятность локализации этих акцепторов УФ-излучения на мембране в непосредственной близости к исследуемому белку. Вместе с тем, учитывая то обстоятельство, что хромофорные группы мембран (порфирины, флавины, нуклеотиды) выступают в качестве сенсибилизаторов пероксидного окисления липидов, можно предположить, что в процесс модификации Na+, К+-АТФазы вносят вклад преимущественно вышеуказанные компоненты эритроцитарных мембран, а также фотосеисибилизированное ими пероксидное окисление липидов.
При УФ-облучении выделенных микросом мозга крыс обнаруживается корреляция между снижением активности Na+, К+-АТФазы и пероксидным окислением липидов, оцениваемым по содержанию полиненасыщенных жирных кислот и накоплению малонового диальдегида (J. Jamme et al., 1995). Подавление активности Na+, К+-АТФазы при облучении микросом УФ-светом снижалось тушителем свободных радикалов — тиомочевиной и не изменялось в присутствии “защитника” тиолов — дитиотреитола. Авторы считают, что эффект ПОЛ опосредуется нарушением целостности мембран, а не структурными изменениями самого фермента.В 1967 г. Liberman показал, что УФ-излучение (255—285 нм) ингибирует Na+, К+-АТФазнук> активность нервных волокон краба. Видимый свет ингибировал Na+, К+-АТФазу различных типов клеток, сенсибилизируемых метиленовым синим, бенгальским розовым или протопорфирином, благодаря фотодинамическому эффекту. Однако чувствительность Na1', К'-АТФазы к видимому свету в отсутствие экзогенных фотосенсибилизаторов не показана. Воздействие излучения аргонового лазера (488 нм, 0,1 Вт/см2) индуцировало ингибирование Na+, К+-АТФазы из мозга крыс дозозависимым образом.
В состав а- (более гидрофобна) и ]3-субъединиц молекул этого белка входят некоторые фосфолипиды, среди которых преобладают фосфатидилэтаноламин, фосфатидилхолин, сфингомиелин. Вероятно, модификация липидного бислоя при облучении мембран затрагивает конформационное состояние не только пограничных (аннулярных) липидов АТФазы, но и непосредственно фосфолипидных компонентов ее а-субъединицы, что приводит к изменению структуры активного центра фермента и нарушению его функциональных свойств.
При изучении УФ-индуцированных изменений ферментативной активности Na+, К+-АТФазы эритроцитарных мембран, модифицированных фосфолипазой D, было установлено, что предварительная обработка мембран фосфолипазой (1О'Й моль/л, pH 5,6; 0,3 моль/л СаС12) приводит к увеличению уровня этого параметра на 33 % по сравнению с контрольным образцом (100 %).
Фосфолипаза D, как уже отмечалось, катализирует отщепление гидрофильного спирта от фосфоглицеридов и сфинго- миелинов. В результате отщепления полярных головок от молекул фосфолипидов могут нарушаться электростатические взаимодействия мембраносвязанных ферментов с молекулами окружающих их фосфолипидов. A. Johannsson et al. (1981) показали, что увеличение или уменьшение толщины липидного бислоя может влиять на активность Na+, Кн-АТФазы за счет перемещения полярных головок фосфолипидов, обращенных к ферментному белку. По-видимому, гидролиз фосфолипидов мембраны, в том числе и аннулярных для Na+, К+-АТФазы, вызывает конфор- мационные изменения молекул фермента, затрагивающие его активный центр, что находит отражение в увеличении уровня активности исследуемого белка.На рис. 48 показаны УФ-индуцированные изменения функциональной активности Na+, К+-АТФазы эритроцитарных мембран, обработанных фосфолипазой D. После модификации фосфолипидов мембраны фоточувствительность связанного фермента существенно изменяется. Воздействие УФ-света на обработанные фосфолипазой мембраны приводит к нарушениям в функционировании Na+, К+-АТФазы, отличающимся по направленности от таковых при облучении интактных мембран, т. е. происходит фотоактивация модифицированного фермента. Следовательно, обработка липидной фазы фосфолипазой нивелирует протекание процессов, обусловливающих фотоинактивацию Na+,
К+-АТФазы в УФ-облученных интактных мембранах, и повышает фотостабильность фермента.
Изменения в функционировании мембраносвязанной Na+, К+-АТФазы в УФ-облученных мембранах, предварительно модифицированных фосфолипазой D, являются, по-видимому, результатом изменения конформационного состояния аннулярных липидов, нарушения белок-липидных взаимодействий, ответственных за проявление каталитической активности фермента.
Обобщая результаты вышеописанных исследований по изучению УФ-индуцированных изменений функциональной активности Na+, К+-АТФазы и АХЭ эритроцитарных мембран, обработанных фосфолипазой D, можно заключить, что в присутствии последней проявляется различная фоточувствительность этих ферментов, обусловленная нарушениями их нативной конформации вследствие модификации белок-липидных взаимодействий, а также влияния фотохимических продуктов молекул мембранных фосфолипидов.
Такие различия в “ответной реакции” двух мембранных белков на воздействие физико-химических факторов (УФ-излуче- ние, действие фосфолипаз) связаны с различиями в их локализации на мембране, в расположении их активных центров, в характере взаимодействия с “ближайшими” липидными молекулами и их химическим строением. Однако экспериментальные данные, касающиеся возможности модуляции фоточувствительности мембранных ферментов, — Na+, К+-АТФазы и ацетилхолинэстеразы, возможно, взаиморегулируемых субстратами их реакций, свидетельствуют в пользу представлений об общности основных регуляторных механизмов функционирования для многих векторных ферментов биомембран.
4.3.
Еще по теме УФ-индуцированные изменения функциональных свойств мем-брапосвязанной Na+, КР-АТФазы в присутствии фосфолипазы D:
- Структура, функциональные и некоторые физикохимические свойства Na+, К[1] АТФазы
- Фоточувствителъностъ ацетилхолинэстеразы эритроцитарных мембран в присутствии фосфолипазы D и аскорбиновой кислоты
- Функциональная активность мембраносвязапной ацетилхолинэстеразы после УФ-облучения в присутствии бензилового спирта и конканавалина А
- Глубинные свойства и экстралингвистические параметры функциональных стилей.
- Нарушение структурно-функциональных свойств клеточных мембран
- Структура, функциональные и некоторые физикохимические свойства ацетилхолинэстеразы
- Изменения в молекуле и биологические свойства
- Влияние изменения свойств строительных материалов во времени
- Окисление, индуцированное ионами металлов
- Генетические варианты, связанные с изменением свойств эритроцитов и обеспечивающие устойчивость к возбудителям малярии.