Функциональная активность мембраносвязапной ацетилхолинэстеразы после УФ-облучения в присутствии бензилового спирта и конканавалина А
Значительный вклад в решение проблемы, касающейся изучения структурного состояния, локализации, особенностей функционирования отдельных компонентов в составе надмолекулярного комплекса, характера их взаимодействия с ближайшим молекулярным окружением, могут внести методические подходы, связанные с модификацией тех или иных структурных элементов мембран.
В качестве модифицирующих агентов, применяемых как для исследования свойств интактных мембран, так и для регуляции метаболических процессов, осуществляющихся с их участием, активно используются разнообразные естественные и синтетические соединения, ферменты, физические факторы — УФ-излучение и температура.Информативным тестом для оценки нативного состояния эритроцитарной мембраны, а также исследования структурных перестроек и изменений в функционировании ее основных компонентов — липидов и белков, индуцированных воздействием целого ряда физико-химических агентов, является определение функциональной активности конформационного маркера мембраны — ацетилхолинэстеразы (см. раздел 1.2.5).
На рис. 32 показаны изменения функциональной активности мембраносвязанной АХЭ после облучения мембран эритроцитов человека УФ-светом в интервалах длин волн 240—390 нм и 300— 400 нм. Видно, что при воздействии УФ-излучения (240—390 нм)
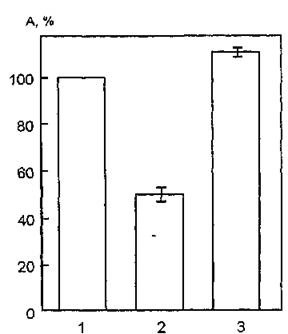
Рис. 32. Функциональная активность ацетилхолинэстеразы мембран эритроцитов человека, модифицированных воздействием УФ-излучения: 1 — контроль; 2 — УФ-облучение в интервале длин волн 300—400 нм; 3 — УФ-облучение в интервале длин волн 240—390 нм. Каталитическую активность регистрировали по методу Хестрина, основанному на колориметрическом определении концентрации ацетилхолина
на суспензию эритроцитарных мембран активность исследуемого фермента статистически достоверно возрастает на 12 %.
Облучение мембран длинноволновым УФ-светом приводит к снижению функциональной активности АХЭ на 50 %. Таким образом, фоточувствительность мембранной АХЭ существенно зависит от спектрального состава УФ-света, а значит, она определяется ролью различных хромофоров мембран при их облучении в указанных диапазонах длин волн.УФ-излучение в интервале длин волн 240—390 нм эффективно поглощается такими структурными компонентами эритроцитарной мембраны, как полиненасыщенные жирные кислоты фосфолипидов, а также ароматические и серосодержащие остатки интегральных белков. Необходимо отметить, что мембранные эффекты УФ-облучения в значительной степени вызываются пероксидным окислением липидов и лишь частично обусловлены фотохимическими превращениями белков. Следовательно, поглощение УФ-излучения в интервале длин волн 240—390 нм указанными выше хромофорами эритроцитарных мембран индуцирует такие структурные перестройки липидного бислоя и интегральных белков, которые, в свою очередь, затрагивают конформационное состояние АХЭ и приводят к увеличению ее функциональной активности.
И. Д. Болотовским и соавт. (1978), С. В. Коневым и И. Д. Болотовским (1979) при изучении влияния структурного состояния липидной фазы мембран эритроцитов на эффективность фо
тохимической модификации АХЭ, связанной с мембраной, обработанной фосфолипазами, а также обедненной холестерином, сделано заключение о важной нефотохимической роли липидной фазы мембран в определении характера и эффективности действия УФ-света на активность фермента.
Учитывая эти данные, можно констатировать, что ПФОЛ, индуцированное УФ-излучением в интервале длин волн 240— 390 нм, не приводит к фотодеструкции мембранной АХЭ, а посредством изменения контролирующих конформацию фермента белок-белковых и белок-липидных взаимодействий способствует более эффективному протеканию каталитической реакции. На наш взгляд, интересным представляется тот факт, что облучение мембран эритроцитов длинноволновым УФ-светом индуцирует резкое снижение каталитической активности АХЭ.
Хромофорами УФ-света в данных условиях эксперимента являются различные (восстановленные) пиридиннуклеотиды, флавины, железопорфи- рины. Итак, ингибирование мембранного фермента в указанном случае может быть обусловлено фотохимическими превращениями вышеназванных хромофоров УФ-излучения. Не исключена вероятность локализации этих акцепторов УФ-света на мембране в непосредственной близости к исследуемому белку. Вместе с тем учитывая то обстоятельство, что хромофорные группы мембран (порфирины, флавины, нуклеотиды) выступают в качестве сенсибилизаторов ПФОЛ, можно предположить, что в процессы модификации АХЭ вносят вклад преимущественно фотохимические превращения указанных компонентов биомембран, а также фотосеисибилизированное ими пероксидное окисление липидов.На рис. 33 представлены результаты исследования ферментативной активности мембранной АХЭ в присутствии конканава- лина А из канавалии мечевидной (Concanavalia ensiformis) в концентрациях 0,4 и 0,8 моль/л. Конканавалин А относится к классу лектинов, связывание которых с поверхностью плазматической мембраны клеток имеет самые разнообразные последствия, например, вызывает изменение в расположении поверхностных белков и гликопротеинов, физическом состоянии липидов мембран, проницаемости их для различных веществ и активности мембранных ферментов (см. раздел 1.3). Конканавалин А, избирательно модифицируя структурно-функциональное состояние интегральных мембранных белков, способен изменять и фоточувствительность АХЭ.
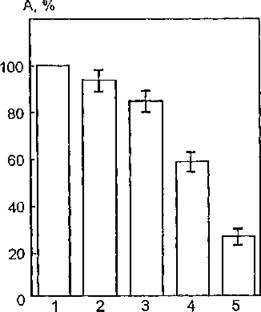
Рис. 33. Каталитическая активносте ацетилхолинэстеразы мембран эритроцитов человека в присутствии конканавалина: 1 — контроль; 2 — конканавалин в концентрации 0,4 моль/л, соотношение суспензии мембран и конка- навалина 1:0,05; 3 — конканавалин в концентрации 0,4 моль/л, соотношение 1:0,2; 4 — конканавалин в концентрации 0,4 моль/л, соотношение 1:1; 5 — конканавалин в концентрации 0,8 моль/л, соотношение 1:1
Из рис, 33 видно, что обработка мембран эритроцитов указанным модифицирующим агентом: вызывает снижение функциональной активности фермента.
При использовании объемных соотношений суспензии мембран и конканавалина 1:0,05 не зарегистрированы статистически достоверные различия величин активности мембранной АХЭ в нативном состоянии и в присутствии лектина. С увеличением этих соотношений до 1:0,2 и 1:1 активность фермента статистически достоверно снижается по сравнению с контролем на 16 и 42 % соответственно. Применение конканавалина в концентрации 0,8 моль/л и соотношении суспензии мембран и экзогенного модификатора 1:1 вызывает ингибирование мембранной АХЭ на 75 %.Основные структурные особенности биологической мембраны и, следовательно, ее функциональная динамичность определяются свойствами липидного бислоя, существенно влияющего на подвижность белковых молекул и их ассоциацию в мембране.
Известно, что бензиловый спирт, взаимодействуя преимущественно с гидрофобными участками липидной фазы, увеличивает подвижность последней, а также изменяет подвижность ее отдельных компонентов.
На рис. 34 показаны изменения функциональной активности мембраносвязанной АХЭ в присутствии бензилового спирта. Инкубирование суспензии мембран с этим модификатором приводит к снижению каталитической активности исследуемого фермента на 16—18 % по сравнению с контролем. Увеличение объема добавляемого спирта до 1 мл не вызывает усиления его ингибирующего действия по отношению к активности молекул АХЭ. Таким образом, увеличение подвижности липидной фазы с помощью бензилового спирта и, тем самым, модификация ближайшего окружения мембраносвязанной АХЭ — фосфатидилсерина индуцирует снижение ее ферментативной активности.
На рис. 35 представлены данные, полученные при исследовании функциональной активности АХЭ мембран эритроцитов, УФ- облученных в интервале длин волн 300—400 нм в присутствии конканавалина А. УФ-облучение интактных мембран эритроцитов индуцирует снижение каталитической активности АХЭ на 50 %, обусловленное, по всей вероятности, поглощением энергии УФ-света пиридиннуклеотидами, флавинами, железопорфири- нами, витаминами, входящими в состав мембраны.
При воздействии УФ-излучения на суспензию мембран эритроцитов в присутствии конканавалина А функциональная активность мембраносвязанной АХЭ возрастает на 45 % по отношению к исследуемому параметру при облучении нативных мембран. Конканавалин А характеризуется спектром поглощения с Хвах при 280 нм, что обусловлено светопоглощением остатков ароматических аминокислот, входящих в состав его молекулы. Следовательно, изменение фоточувствительности мембранной АХЭ в присутствии лектина может быть связано с влиянием на ее структурное состояние продуктов фотохимических превращений нуклеотидов, коферментов, железопорфиринов и витаминов, интенсивно поглощающих свет в данной области спектра.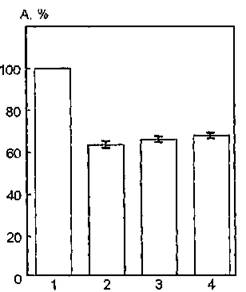
Рис. 34. Функциональная активность ацетилхолинэстеразы мембран эритроцитов человека, модифицированных 2 %-ным раствором бензилового спирта: 1 — контроль; 2 — соотношение суспензии мембран и бензилового спирта 1:0,05; 3 — соотношение этих компонентов 1:0,2; 4 — соотношение 1:1
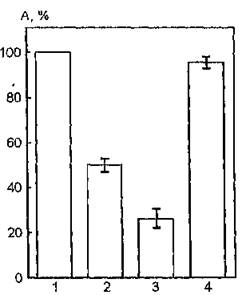
Рис. 35. Ферментативная активность ацетилхолинэстеразы мембран эритроцитов человека, облученных УФ-светом в области 300—400 нм в присутствии конканавалина А: 1 — контроль; 2 — активность УФ- облученной АХЭ; 3 — активность АХЭ в присутствии конканавалина; '4 —активность УФ-облученной АХЭ в присутствии конканавалина
На рис. 36 показаны изменения ферментативной активности АХЭ, модифицированной воздействием УФ-излучения (300— 400 нм) в присутствии бензилового спирта. Из анализа рисунка следует, что облучение суспензии эритроцитарных мембран в присутствии указанного химического агента индуцирует резкое возрастание каталитической активности белка: 210 % по сравнению с ее уровнем при УФ-облучении интактных мембран (50 %).
Наблюдаемый эффект активации мембранной АХЭ может быть связан, по всей вероятности, со структурными перестройками молекул ближайшего липидного окружения фермента, обусловленными воздействием на него бензилового спирта и, прежде всего, его фотохимических продуктов.
Вследствие разрыхления липидной фазы мембраны, модифицированной бензиновым спиртом, и увеличения подвижности ее отдельных компонентов возможно ослабление связей АХЭ с молекулами, находящимися в непосредственном контакте с ней, а именно фосфатидилсерина. Результатом этих процессов, по-видимому, являются конформационные превращения всей молекулы АХЭ и демаскирование ее активного центра на поверхности эритроцитарной мембраны, проявляющиеся в резком возрастании функциональной активности фермента.При УФ-облучении мембран в диапазоне длин волн 240— 390 нм в присутствии указанных экзогенных модификаторов наблюдаются противоположные по направлению изменения функционирования АХЭ.
На рис. 37 представлены результаты исследования каталитической активности мембраносвязанной АХЭ при воздействии УФ-излучения (240—390 нм) на суспензию мембран эритроцитов в присутствии конканавалина А (фитогемагглютинина). Из
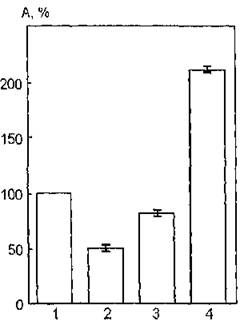
Рис. 36. Каталитическая активность ацетилхолинэстеразы мембран эритроцитов человека, облученных УФ-светом в области 300—400 нм в присутствии 2 % -ного бензилового спирта: 1 — контроль; 2 — активность УФ-облученной АХЭ; 3 — активность АХЭ в присутствии бензилового спирта; 4 —- активность УФ-облученной АХЭ в присутствии бензилового спирта
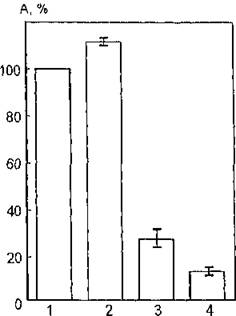
Рис. 37. Функциональная активность ацетилхолинэстеразы мембран эритроцитов человека, облученных УФ-светом в области 240—390 нм в присутствии конканавалина: 1 — контроль; 2 — активность УФ-облученной АХЭ; 3 — активность АХЭ в присутствии конканавалина; 4 — активность УФ-облученной АХЭ в присутствии конканавалина
анализа рисунка вытекает, что УФ-облучение мембран эритроцитов в его присутствии индуцирует значительное снижение активности этого фермента до 12 % (против 112 % при облучении интактных мембран). Вероятно, ингибирование мембраносвязанной АХЭ в данном случае происходит за счет конформационных изменений молекул фермента, обусловленных воздействием на последние, а также на молекулы интегральных белков продуктов фотолиза конканавалина и приводящих, по-видимому, к стери- ческому экранированию активного центра. Необходимо отметить, что УФ-облучение буферного раствора (pH 7,6) конканавалина А вызывает статистически достоверное снижение величины оптической плотности в максимуме поглощения при 280 нм, связанное либо с разрушением части хромофоров УФ-света лектина — ароматических аминокислотных остатков, либо с их экранирова-
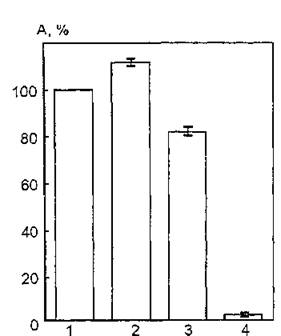
Рис. 38. Ферментативная активность ацетилхолинэстеразы мембран эритроцитов человека, облученных УФ-светом в области 240—390 нм в присутствии 2%-ного бензилового спирта: 1 — контроль; 2 — активность УФ-облученной АХЭ; 3 — активность АХЭ в присутствии бензилового спирта; 4 — активность УФ-облученной АХЭ в присутствии бензилового спирта»
нием на поверхности белковой глобулы. Однако не исключена вероятность и прямого воздействия УФ-излучения на АХЭ.
При УФ-облучении (240—390 нм) эритроцитарных мембран в присутствии бензилового спирта обнаруживается практически полное ингибирование мембранной АХЭ (рис. 38). По-видимому, наблюдаемый эффект может быть обусловлен реализацией нескольких параллельно протекающих процессов, индуцированных воздействием УФ-излучения на эритроцитарные мембраны и приводящих к экранированию активного центра:
1) возможная интенсификация ПФОЛ вследствие увеличения подвижности липидной фазы мембран под влиянием бензилового спирта, образование продуктов ПФОЛ, влияющих на конформацию АХЭ и ее активного центра непосредственно или опосредованно через взаимодействие с интегральными белками;
2) фотопревращения интегральных белков мембраны, поглощающих свет в данной области спектра, в том числе и самого фермента (однако в незначительной степени);
3) поглощение квантов УФ-излучения с длинами волн 330— 370 нм пиридиннуклеотидами, флавинами, коферментами, же- лезопорфиринами; фотосенсибилизация ими процессов ПФОЛ; непосредственное влияние их фотохимических продуктов на структурное состояние АХЭ и ее активного центра.
Таким образом, путем модификации отдельных структурных компонентов эритроцитарных мембран различными химически-
ми агентами и выбора условий их УФ-облучения в зависимости от природы и содержания хромофорных групп можно существенно изменять фоточувствительность мембранных белков, в том числе АХЭ.
4.2.1.
Еще по теме Функциональная активность мембраносвязапной ацетилхолинэстеразы после УФ-облучения в присутствии бензилового спирта и конканавалина А:
- Фоточувствителъностъ ацетилхолинэстеразы эритроцитарных мембран в присутствии фосфолипазы D и аскорбиновой кислоты
- Структура, функциональные и некоторые физикохимические свойства ацетилхолинэстеразы
- УФ-индуцированные изменения функциональных свойств мем-брапосвязанной Na+, КР-АТФазы в присутствии фосфолипазы D
- Механизмы регулирования функциональной активности ферментов и ферментных систем в клетке
- Динамика индекса активности и распространенности процесса mLoSSI при ограниченной склеродермии после различных методов лечения
- 41. Этиловый спирт. Отравления. Привыкание и зависимость. Медицинское применение.
- 7. Концепция А.Р. Лурии о трех основных структурно-функциональных блоках. 8. Специфика вклада структурно-функциональных блоков в осуществление высших психических функций.
- №50. Функциональные ряды, область сходимости функциональных рядов. Равномерная сходимость. Теорема Вейерштрасса.
- Статья 63 Судебное разбирательство в присутствии обвиняемого
- Глава 14. Сам факт присутствия других
- Присутствие (и отсутствие)
- Инфракрасные датчики движения и присутствия
- Присутствие полномочных лиц и их представителей при таможенном оформлении (ст. 128)
- Дети наследовали не только после отца, но и после матери.
- 31. Разграничение правонарушений по присутствию и отсутствию виновности учинителя
- 25. Разграничение правонарушений по присутствию и отсутствию различных стадий развития
- 29. Разграничение правонарушений по присутствию и отсутствию в них безнравственного характера
- Англичане, ваххабиты и египтяне на Бахрейне. Формы английского колониального присутствия {X!X в.)
- При взгляде на название данной главы читатель, вероятно, испытывает некоторое удивление: почему в книге о профессиональных навыках юриста присутствует рассказ об эволюции правозащитного движения?