Окисление, индуцированное ионами металлов
Характер изменения состава ЛНП при окислении [584], защитная эффективность антиоксидантов [587], зависимость от кислорода [1563] указывают на свободнорадикальный механизм этого процесса, окислению в первую очередь подвержены ненасыщенные липиды и жирные кислоты [703, 1359].
Ионы металлов переменной валентности, разлагая органические перекиси, выступают инициаторами окисления ЛНП. Инкуба-ция in vitro частиц ЛНП с низкими концентрациями (1—20 мкМ) Cu или Fe приводит к изменению их состава (см. табл. 5), физических и биологических свойств (см. табл. 6); кинетические параметры этих изменений в настоящее время подробно изучены [2, 41, 171, 589]. В физиологических условиях при наличии в среде растворенного кислорода концентрация пероксильных радикалов [LO'] значительно больше концентрации [L'], поэтому в отсутствие антиоксидантов ингибирование окисления идет главным образом посредством диспропорционирования радикалов LО'. Если принять скорость зарождения радикалов в среде за V, то для стационарного процесса (d[LO2]/dt = 0) получаем:
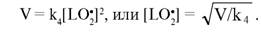
В процессе окисления нарабатываются органические перекиси. Важным показателем, характеризующим цепное окисление, является длина цепи окисления (ДСО), которая показывает, сколько молекул перекисей образуется на один акт зарождения радикалов: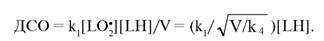
Оценки показывают, что при окислении ненасыщенных жирных кислот в нормальных условиях ДСО находится в пределах от 1 до 200. Так, при окислении лино- левой кислоты ДСО = 26 [1359]; это означает, что каждый зародившийся радикал дает начало 26 молекулам перекисей.
При наличии в среде антиоксидантов ингибирование окисления преимущественно идет в реакции LO' с молекулами антиоксидантов. В этом случае для стационарного процесса имеем: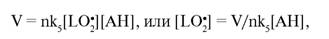
где n — количество радикалов, ингибируемых молекулой антиоксиданта (для α-токо- ферола n = 2); в этом случае ДСО = kJLH]/nk5[AH].
Как показывают эксперименты, добавление 1 молекулы α-токоферола на 1000 молекул ненасыщенных жирных кислот снижает ДСО в 10—500 раз. Такая зависимость от антиоксидантов приводит к тому, что окисление ЛНП, индуцированное ионами металлов, носит фазовый характер (рис. 10).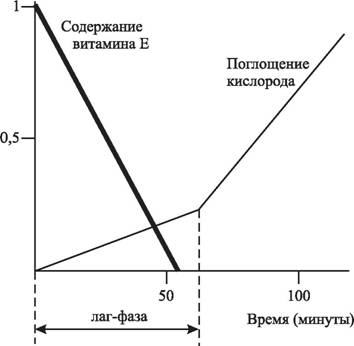
Рис. 10. Кинетика изменения окисления ЛНП в присутствии 2,2'-азобис(2-амидино- пропан)дигидрохлорида [585]
В начальной фазе (лаг-фаза), продолжительность которой определяется выражением T = n[AH]/V, происходит окисление антиоксидантов и незначительно возрастает содержание гидроперекисей и других продуктов ПОЛ. Введение экзогенных антиоксидантов увеличивает продолжительность лаг-фазы [586]. После истощения жирорастворимых антиоксидантов начинается эффективное окисление жирных кислот, что сопровождается усилением поглощения кислорода, значительно возрастает содержание гидроперекисей и вторичных продуктов ПОЛ [585]. Связывание продуктов ПОЛ с апо В приводит к его химической модификации, что усиливает поглощение ЛНП макрофагами [824, 1564].
Активность окисления в присутствии ионов меди существенно зависит от состава ЛНП, и прежде всего — от наличия ненасыщенных жирных кислот, свободного холестерина и убихинона [919]. Ионы меди более эффективны в окислении ЛНП, чем ионы железа, при этом другие ионы металлов переменной валентности значительно менее эффективны [937]. В отличие от Fe-индуцированного окисления, которое требует присутствия в среде восстановителей (например, О2), Cu-индуцированное окисление ЛНП может протекать в отсутствие специальных восстановителей, так как восстанавливать ионы меди может α-токоферол:
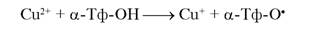
С помощью специфических антител, полученных к окисленным in vitro ЛНП, повышенное содержание таких липопротеинов обнаруживается в атеросклеротически поврежденных сосудах [772] и крови больных атеросклерозом [1447].
У людей с повышенным содержанием ионов Cu2+ в сыворотке показывают утолщение стенки сонной артерии [1446], а эпидемиологические исследования обнаруживают прямую взаимосвязь между содержанием в сыворотке ионов меди [496], уровнем запасенного железа [1447], с одной стороны, и содержанием продуктов радикального окисления липидов и риском развития коронарной болезни сердца — с другой. В экспериментах на кроликах было показано, что введение декстрана железа с одновременной индукцией гиперхолестеринемии (рацион с 0,5 % холестерина) усиливает атеросклеротическое поражение сосудов, при этом повышается содержание гидроперекисей в гомогенатах тканей и наблюдается накопление железа в артериальной стенке [270]. Эти результаты позволяют сделать вывод о возможности окисления циркулирующих ЛНП ионами металлов переменной валентности во внеклеточных средах, что косвенно подтверждается наличием окисленных ЛНП в плазме здоровых людей [285].В организме железо связывается с трансферрином и ферритином, а медь — с церулоплазмином и альбумином, поэтому неясно, каково происхождение данных ионов и могут ли они участвовать в окислении ЛНП in vivo. При введении окисленных ЛНП в кровь животных они быстро удаляются клетками печени (за 1 минуту концентрация меченых ЛНП снижается на 70 % [210]); поэтому неизвестно, происходит ли окислительная модификация липопротеинов в сыворотке. Так как даже незначительное (несколько процентов) присутствие сыворотки в среде ингибирует окислительную модификацию ЛНП, индуцированную ионами металлов или клетками, то многие исследователи считают, что в организме окисление возможно только в области атеросклеротической бляшки [641, 985, 984]. Одним из аспектов данной проблемы является соответствие широко распространенной экспериментальной модели окисления ЛНП в присутствии ионов Cu2+ и Fе2+ ситуации, происходящей в живом организме.
В гомогенатах атеросклеротических бляшек артерий человека было измерено содержание ионов железа и меди.
Общее содержание ионов железа находилось в пределах 2,9—56,3 мкМ, при этом каталитически активный пул составлял около 10 % (среднее значение — 1,14 мкМ), содержание ионов меди составляло от 0,2 до 28,6 мкМ [1536]. Гомогенаты атеросклеротических бляшек в 2—3 раза усиливали ПОЛ в микросомах печени в присутствии аскорбиновой кислоты, при добавлении перекиси водорода наблюдалось возникновение ОН-радикала, выявлялось также образование супероксидного аниона, что свидетельствует о наличии каталитически активных ионов металлов пере-менной валентности [1536, 1590]. Кроме того, было показано, что экстракты из бляшек могут прямо окислять ЛНП [766].
В экспериментах in vitro было установлено, что источниками каталитически активных ионов железа может выступать ферритин, гемин и трансферрин, а ионов меди — церулоплазмин. Так, было показано, что ферритин является одним из основных источников каталитически активного железа в атеросклеротической бляшке [41, 332]. В клетках также существует микросомальный пул негемового железа, высвобождение которого также может индуцировать процессы ПОЛ [1154]. В атеросклеротических бляшках обнаружено высвобождение каталитически активного железа из трансферри- на [965]. Разлагать органические перекиси и окислять ЛНП могут не только свободные ионы железа, но и входящие в состав гема, в частности, гемин [295], гемоглобин разрушенных эритроцитов [1261] или цитохром с, высвобождающийся из поврежденных митохондрий [1369]. Кроме того, при инкубации с Н2О2 из гемовых белков высвобождаются каталитически активные ионы железа [699, 715].
Около 90 % ионов меди в сыворотке человека связано с церулоплазмином, главным медьсодержащим ферментом. В экспериментах по индуцированному ионами железа и меди окислению липидных субстратов было показано, что церулоплазмин эффективно ингибирует ПОЛ [698]. Вместе с тем высказывается предположение, что в организме человека церулоплазмин является скорее прооксидантом, окисляя ЛНП, нежели антиоксидантом [571]. Это связано с тем, что в концентрациях больше 100 мкг/мл церулоплазмин индуцирует окислительную модификацию ЛНП: эффективность накопления в частицах ЛНП ТБК-РП, гидроперекисей и диеновых конъюгатов при инкубации со 100 мкг/мл церулоплазмина была эквивалентна значениям, полученным при инкубации с 0,7 мкМ С^О4; удаление из молекулы одного атома меди отменяет способность к окислению [571]. На необходимость более тщательного изучения роли церулоплазмина в окислении ЛНП указывает тот факт, что это соединение может накапливаться в стенке артерий за счет специфического рецепторного связывания [1571] и за счет клеточного синтеза в процессе воспалительного повреждения [608].