Структура, функциональные и некоторые физикохимические свойства ацетилхолинэстеразы
Один из важнейших биохимических механизмов, лежащих в основе деятельности нервной системы, — это химический процесс, связанный с биосинтезом, выделением и распадом ацетилхолина.
Скорость ферментативного гидролиза последнего играет ключевую роль в функционировании нервной системы. Изменения в кинетике ферментативного разрушения ацетилхолина приводят к серьезным нарушениям функций этой системы. Одна из областей фармакологии разрабатывает способы лечения заболеваний нервной системы путем применения лекарственных средств, влияющих на кинетические параметры гидролиза ацетилхолина холинэстеразами.После обнаружения Дейлом в 1914 г. в крови фермента, катализирующего процесс расщепления ацетилхолина, было показано широкое распространение этого фермента в различных тканях животных. В 40-е гг. было высказано предположение о существовании двух основных типов ферментов, гидролизующих эфиры холина, — “истинных холинэстераз” и “псевдохолинэстераз”. Эти ферменты относятся к сериновым гидролазам и соответствуют двум группам биологических катализаторов: ацетил- холинэстеразе (ацетилгидролаза ацетилхолина, КФ 3.1.1.7) и холинэстеразе (ацилгидролаза ацилхолинов, КФ 3.1.1.8). Холинэстеразы наряду с гидролизом ацетилхолина могут участвовать также в межклеточных взаимодействиях. С этим связана специфическая экспрессия холинэстераз в эмбриогенезе в определенных структурах до формирования холинэргических синапсов, а также обнаружение их в нехолинэргических тканях.
Ферментативное расщепление ацетилхолина представляет собой реакцию гидролиза, протекающую с образованием уксусной кислоты и холина:
(CH3)3N+CH2CH2OC(0)CH3 + Н2о -4 (СН8)3№СН2СН2ОН + СН3С00Н
Основное назначение холинэстеразы нервной ткани — быстрый гидролиз выделяющегося ацетилхолина, без которого невозможна передача нервных импульсов.
Обнаружено, что ферментативный гидролиз ацетилхолина может осуществляться не только нервной, но и другими тканями: присутствие холинэстеразы было выявлено в сыворотке и эритроцитах крови, в мышечной ткани, печени, поджелудочной железе. Считают, что холинэстераза эритроцитов играет важную роль в клеточной проницаемости. Холинэстераза крови и тканей рассматривается как своеобразный защитный (“аварийный”) фермент на случай значительных выходов в кровяное русло ацетилхолина при перевозбуждении нервной системы.К ацетилхолинэстеразам относятся ферменты нервной ткани и эритроцитов, а к холинэстеразам — ферменты сыворотки крови, печени, поджелудочной железы и других органов. Ацетилхо- линэстераза катализирует гидролиз ацетилхолина и ацетил- р-метилхолина и не влияет на гидролиз бензоилхолина и бути- рилхолина. Холинэстераза катализирует гидролиз бутирилхоли- на, ацетилхолина, бензоилхолина и пропионилхолина, но не действует на ацетил-Р-метилхолин. Активность ацетилхолинэстера- зы характеризуется отчетливым максимумом при концентрации ацетилхолина ~ 2-Ю’4 моль/л и тормозится при ее увеличении, в то время как на активность холинэстеразы не влияет избыток ацетилхолина. Оптимум pH-действия составляет для аце- тилхолинэстеразы 7,5—8,0, а для холинэстеразы — 8,5 (рис. 12).
В каталитическом действии холинэстераз обоего типа принимает непосредственное участие гидроксильная группа одного из остатков серина, расположенного на активной поверхности фермента. Однако такой гидроксил должен быть специфически активирован, чтобы приобрести способность к участию в каталитическом действии. Эта активация может быть осуществлена путем взаимодействия с группировкой белковой молекулы, характеризующейся величиной рК 5,8—7,0. Предполагают, что такой группировкой является остаток гистидина. Доказано наличие в холинэстеразах обоего типа анионной группировки, несущей единичный отрицательный заряд и взаимодействующей с катионной группировкой субстрата — ацетилхолина, а также катионсодержащих ингибиторов.
В активном центре холинэстеразы вблизи серина находится остаток глутаминовой кислоты. Проведены расчеты электростатического потенциала и электрического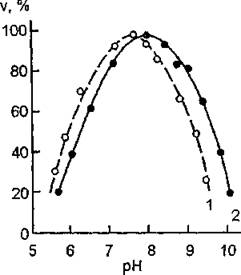
Рис. 12. Зависимость скорости гидролиза ацетилхолина от pH для ацетилхолинэстеразы Torpedo marxnorato (У) и холинэстеразы сыворотки крови (2}
•поля вокруг наиболее важного в каталитическом отношении “горла” ацетилхолинэстеразы, включающего серин, гистидин и глутамат. Из результатов исследования pH-зависимости действия холинэстераз следует, что их активность связана с функциями группировки, рК которой составляет 8,5— 10,0. К таким группировкам может быть отнесен гидроксил тирозина. В образовании фермент-суб- стратного комплекса ацетилхолинэстеразы участвует в качестве обязательных не менее трех связей. Во всех современных схемах механизма действия холинэстераз фигурируют: гидроксил серина, связанный с имидазолом гистидина, — в качестве постулированной Уилсоном нуклеофильной группировки эстеразного центра; ионизированная карбоксильная группа — в качестве анионного центра; гидроксил тирозина — в качестве кислотной группы эстеразного центра.
Считают, что каталитический процесс (рис. 13) начинается с образования электростатической связи между катионом субстрата и анионной группировкой фермента. Нейтрализация отрицательного заряда одной из групп белковой молекулы в сочетании с взаимодействием трех метильных групп при атоме азота ацетилхолина с окружающими анионный центр группами приводит к существенным изменениям конформации фермента. В результате этого группировки эстеразного центра и соответствующие группы субстрата занимают положение, обеспечивающее установление “наилучшей” комплементарности, а затем образуются связи между углеродом поляризованной карбонильной группы и кислородом гидроксильной группы серина и, возможно, между эфирным кислородом и водородом кислотной группировки (остаток тирозина). Далее происходит переход протона к холину с образованием ацетилированного по гидроксилу фермента и про- тонированного имидазола.
Следующий элементарный акт с участием воды осуществляется после того, как молекула холина покинет анионный центр фермента и восстановится отрицательный заряд анионной группировки. Молекула воды образует связи с карбонильным кислородом и кислородом тирозина, в результате чего происходит переход протона к гидроксилу серина. При этом выделяется второй продукт реакции — уксусная кислота — и регенерируется фермент в исходной конформации.Эксперименты Е. Krejci и соавт. (1991) с направленным мутагенезом показали, что консервативный для эстераз и липаз остаток аспарагиновой кислоты необходим для осуществления ими каталитических функций. По-видимому, электростатические особенности поверхности молекулы ацетилхо- линэстеразы не влияют на скорость катализа, которая не зависит от диффузии молекул субстрата, а стабилизация переходных состояний в каталитическом центре нечувствительна к электростатическим взаимодействиям.
Для выделения ацетилхо- линэстеразы из эритроцитов используют метод экстракции фермента из стромы щелочным буфером в присутствии нейтрального детергента Твин-20 с последующим фракционированием сульфатом аммония и отделением гемоглобина с помощью кальций-фосфатного геля. Молекулярная активность (т.е. число молекул субстрата, претерпевающих превращение на одном активном (каталитическом) центре фермента в 1 мин в условиях насыщения фермента субстратом) ацетилхолинэстеразы составляет - (3—3,5)-105 мин-1 и превышает в 4— 5 раз таковую для холинэстеразы сыворотки крови.
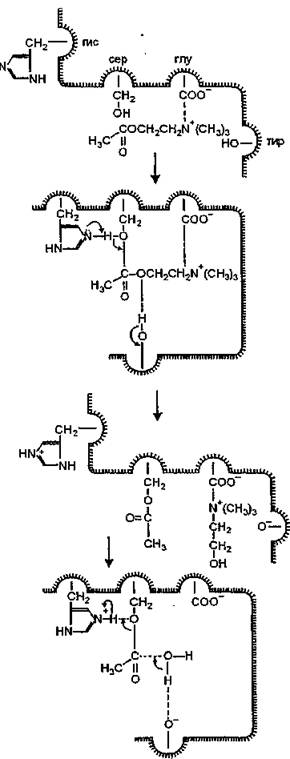
щие. 13. Схема механизма действия холинэстераз
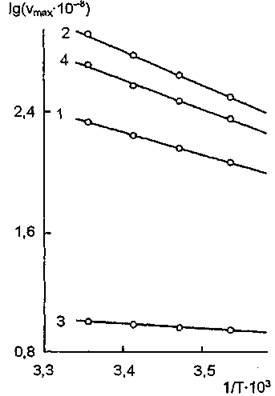
Рис. 14. Зависимость lgv от 1/Т для ферментативного гидролиза эфиров холина при действии холинэстеразы сыворотки крови лошади: 1 — ацетилхолин; 2 — бутирилхо- лин; 3 — ацетил-р-метилхолин; 4 — бензоилхолин
Зависимость lg v от 1/Т для ферментативного гидролиза эфиров холина холинэстеразой сыворотки крови лошади (график Аррениуса) в интервале температур 10—25 °С является линейной (рис.
14). В случае гидролиза ацетил-Р-метилхолина константа Михаэлиса не зависит от температуры, для ацетилхолина и бен- зоилхолина — незначительно растет по мере ее повышения, а для бутирилхолина — существенно увеличивается.Ацетилхолинэстераза эритроцита расположена на внешней поверхности мембраны и составляет 0,2—0,3 % от всего мембранного белка. Она является гликопротеином, в состав которого входят гликано- вые компоненты, содержащие последовательность: этанола- мин — фосфатманноза — глю- козамин — инозитол. Фермент может быть экстрагирован в форме активного липопротеина при обработке мембраны растворами высокой ионной силы, Na-дезоксихолатом или тритоном Х-100. Удаление липида приводит к полной потере активности фермента, а последующее добавление фос- фатидилсерина (липида, с которым он выделяется из мембраны) — к восстановлению ее функциональных свойств.
Ацетилхолинэстераза состоит из двух одинаковых субъединиц с молекулярной массой, равной 70—90 кДа. Каждая субъединица имеет два центра связывания: активный и аллостерический. Активность фермента зависит от ионного состава среды и специфически ингибируется ионами тетраметиламмония, производными карбаминовой кислоты и фосфоорганическими соединениями. Показано, что хлорпирофосметил (ХПМ) является сильным неконкурентным ингибитором ацетилхолинэстеразы из эритроцитов человека. При растворении ацетилхолинэстеразы в тритоне Х-100 ее активность и кинетические свойства не изменяются, а также сохраняется чувствительность к ХПМ. Предполагают, что на участки связывания ацетилхолинэстеразы и ХПМ не влияет гидрофобное окружение мембран эритроцитов. Ацетилхолинэстераза является аллотопным ферментом-, каталитические свойства ее существенно различаются в растворе и в составе биомембран.
Каталитическая активность мембранной ацетилхолинэстеразы находится под контролем структурного состояния липидной фазы эритроцитарной мембраны. Фосфолипазы (А2, С и D) оказывают на мембраны близкое модифицирующее действие, хотя они характеризуются не только различной специфичностью (природой разрываемых в липидах связей), но и пространственной асимметрией действия.
Панкреатическая фосфолипаза А2 и фосфолипаза D гидролизуют липиды, расположенные на обеих сторонах эритроцитарной мембраны, а фосфолипаза С гидролизует фосфолипиды, расположенные на внутренней ее стороне. Модификация липидного бислоя с внутренней стороны мембраны приводит к изменению структурного состояния липидов, а затем и белков, расположенных снаружи. Косвенным свидетельством возможности такой трансмембранной передачи структурного сигнала может служить отрыв ацетилхолинэстеразы от мембраны под влиянием фосфолипазы С (И. Д. Болотовский и соавт., 1987). Обработка любыми фосфолипазами как бы превращает мембраносвязанный фермент в квазисвободный.Следует отметить, что поскольку функциональные свойства ацетилхолинэстеразы существенно зависят от структурного состояния мембраны, то определение уровня ее активности используют в качестве конформационного маркера для оценки различного рода модификаций мембранных компонентов под влиянием физико-химических факторов (см. главу 4).
Кроме того, активность холинэстераз изменяется при многих патологических процессах: заболеваниях печени, почек, остром инфаркте миокарда, онкозаболеваниях, в послеоперационном периоде, при интоксикации фосфорорганическими соединениями, поэтому эта характеристика белковой молекулы может быть использована в энзимодиагностике.
1.3.