ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА ЛАКТАТДЕГИДРОГЕНАЗЫ В КОМПЛЕКСЕ С ЭРИТРОЦИТАРНЫМИ МЕМБРАНАМИ И ИХ КОМПОНЕНТАМИ В ИНТАКТНОМ СОСТОЯНИИ И ПОСЛЕ УФ-ОБЛУЧЕНИЯ
В настоящее время считается общепризнанным, что “классическое” исследование ферментов in vitro без учета их взаимодействия с различными субклеточными структурами не может дать адекватного представления о закономерностях и механизмах их функционирования в составе целой клетки.
Поэтому широко обсуждаются вопросы, связанные с локализацией гликолитических ферментов в клетке (см. раздел 2.3.2) и, в частности, лактатдегидрогеназы.L-лактатдегидрогеназа (Ь-лактат:ЫАВ|'-оксидоредуктаза, КФ 1,1.1.27) — гликолитический фермент, относящийся к классу оксидоредуктаз. Он катализирует обратимое окисление L-лактата до пирувата с использованием в качестве кофермента NAD+.
Лактатдегидрогеназа (ЛДГ) принадлежит к наиболее интенсивно изучаемым белкам, так как она является ключевым ферментом, функционирующим на «развилке» путей аэробного и анаэробного превращения углеводов в тканях.
Молекула ЛДГ представляет собой тетрамер, состоящий из субъединиц двух типов — Н и М (от англ, heart — сердце и muscle — мышца). Молекулярные массы тетрамера и каждой субъединицы составляют 140 и 35 кДа соответственно. Рекомбинация двух типов субъединиц дает различные изоформы фермента: гомотетрамеры Н4 и М4 и гибридные тетрамеры (гетеротетрамеры) Н3М, Н2М2 и НМ5. Это количество изоферментов обусловлено наличием двух генетических локусов, которые кодируют синтез субъединиц М и Н .
Субъединица М синтезируется главным образом в тканях с анаэробным метаболизмом, в то время как субъединица И присутствует в тканях с преобладанием аэробных процессов. В цитозоле герминативных клеток, в частности в сперматозоидах, присутствует ЛДГ-Х, составляющая 80 % активности ЛДГ. Показано, что ЛДГ-Х также является тетрамером и состоит из субъединиц ЛДГ-Х, которые по аминокислотному составу отличаются от Н и М.
Изоферментам ЛДГ присущи разные кинетические характеристики, величины pH, при которых они проявляют максимальную активность, сродство к субстратам и кофакторам. Для определения относительного содержания изоферментов используют ряд физико-химических методов: электрофорез, ионообменную хроматографию, различия в кинетических параметрах отдельных изоферментов, дифференцированную чувствительность к субстратам и ингибиторам, иммунохимические методы.
Полипептидная цепь обеих субъединиц содержит 330 аминокислотных остатков; различия в их последовательности в субъединицах обнаружены на протяжении более чем 25 % длины полипептидной цепи.
В субъединицах фермента выделяют два домена: каталитический и центр связывания кофермента. Аминокислоты, образующие ЫАГГ-связывающий домен, расположены в середине молекулы ЛДГ. Структура названного домена состоит из шести складчатых слоев (ЗА—pF), соединенных между собой ос-спиралями.
Лактатдегидрогеназа связывает лактат или пируват только в присутствии кофермента, причем первым связывается кофермент. NAD+ (NADH) находятся в гидрофобной полости в изогнутой конформации молекулы ЛДГ.
В каталитическом центре ЛДГ находятся полярные группы, участвующие в связывании кофермента и субстратов и катализе: остатки Arg 109, Arg 171, Arg 101, Glu 102, Asn 140, а также существенный остаток His 195, способный играть роль донора- акцептора И1 в ходе реакции. Кроме того, для проявления активности фермента необходима сульфгидрильная группа, одна на субъединицу.
Оптимум pH для реакции восстановления пирувата в лактат — 6,5—7,5, а для обратной реакции — 8,5—9,0. Ингибируют фермент высокие концентрации NAD, фторпируват (необратимо), а-меркаптокислоты (конкурентно по отношению к лактату), сульфгидрильные реагенты. В последнем случае ингибирование предотвращается коферментом и снимается цистином и глутатионом.
Получены сведения о взаимодействии ЛДГ со структурными белками мышц и миофибриллами, субклеточными частицами, искусственными подложками, микросомами и мембранами саркоплазматического ретикулума.
Установлено, что взаимодействие ЛДГ с F-актином и нативным тонким филаментом приводит к уменьшению каталитической активности фермента. При связывании с F-актином происходит увеличение значения константы Михаэлиса для NADH и уменьшение максимальной скорости реакции. Для свободной и связанной форм ЛДГ выполняется уравнение Михаэлиса — Мен- тен. Предполагают, что обратимое взаимодействие изофермента М) ЛДГ с F-актином является специфическим и обусловлено образованием белок-белкового комплекса.
Исследование кинетических параметров ЛДГ в растворе и в суспензии гомогената субклеточных частиц скелетных мышц цыпленка позволило выявить, что изофермент ЛДГ Н4 не связывается субклеточными частицами, ЛДГ М2 полностью и ЛДГ Н2М2 частично находятся в связанном состоянии. Связывание изоферментов М4 и Н2М2 приводит к значительному снижению величины максимальной скорости реакции, а также константы Михаэлиса для пирувата.
При взаимодействии ЛДГ с митохондриальной фракцией и митохондриальным ингибитором из скелетных мышц кролика отмечено снижение активности фермента. Причем зависимость скорости реакции от концентрации субстратов для связанной ЛДГ была негиперболической, а зависимость І/v от 1/[пируват]1/2 или l/[NADH]3/2 была линейной.
Описано связывание ЛДГ М4 с искусственными подложками, карбоксиметилцеллюлозой и декстрансульфатом. Взаимодействие ЛДГ из мышц свиньи с декстрансульфатом приводит к появлению положительной кооперативности по концентрации NADH, не характерной для фермента в растворе.
В. И. Лущак (1991) изучил физико-химические свойства ЛДГ, связанной с микросомальными мембранами из белых мышц шиповатого ската. Исследование кинетических характеристик для мембраносвязанного и изолированного ферментов показало, что константа Михаэлиса для данных форм (состояний) ЛДГ практически одинакова. В то же время данный показатель для NADH у связанного фермента в 4 раза ниже, чем у изолированного.
Цитозольная форма ЛДГ из белых мышц шиповатого ската ингибируется наполовину при концентрации пирувата 14 ммоль/л, а связанная проявляет в этих условиях полную активность. Связанная ЛДГ обладает более высокой стабильностью, чем свободная: перевод фермента в раствор в 2,5—8,0 раза ускоряет процесс его инактивации трипсином. Такие факторы, как pH, ионная сила, субстраты и коферменты, влияют на процесс экстракции ЛДГ из микросом. При щелочных значениях pH удается солюбилизировать 48 % активности фермента, но не менее 20 % ЛДГ остается в мембраносвязанном виде. Степень экстрагируе- мости фермента из микросом возрастает по мере увеличения ионной силы раствора. При нейтральных значениях pH среды NADH экстрагирует 68 % ЛДГ. По-видимому, с микросомами из белых мышц шиповатого ската связано два пула ЛДГ. Фермент одного пула взаимодействует с мембраной за счет электростатических сил, фермент другого пула — за счет гидрофобных взаимодействий. Наличие двух пулов ЛДГ приводит к дополнительной возможности регуляции свойств фермента путем его перераспределения между разными клеточными компартментами,Т. В. Есаковой и М. В. Ивановым (1992) показано, что связывание ЛДГ с мембранами саркоплазматического ретикулума ведет к снижению удельной активности фермента на 60—70 %. Высказано предположение о том, что взаимодействие фермента с мембранами происходит в участках с высоким и низким сродством. Добавление NADH, NAD+, а также повышение ионной силы раствора вызывает солюбилизацию связанного фермента. Снижение активности ЛДГ в связанном состоянии можно объяснить как маскировкой некоторых субъединиц, диффузионными затруднениями при связывании субстратов, конформационными перестройками молекулы ЛДГ, так и изменениями физико-химических свойств среды при переходе фермента из раствора в при- мембранный слой.
Взаимодействие мембран эритроцитов с ЛДГ приводит к снижению ее каталитической активности. На рис. 49 представлена зависимость ферментативной активности мембраносвязанной ЛДГ от величины pH среды инкубирования.
В условиях оптимума pH 7,4 активность ее снижается на 25 %; при значениях pH 6,0 и 4,0 — соответственно на 33 и 46 %. Сдвиг pH в щелочную область приводит к уменьшению эффекта ингибирования ЛДГ. При pH 9,0 активности свободного и связанного ферментов статистически достоверно не отличаются.По-видимому, с увеличением отрицательного заряда молекулы белка вероятность образования его комплекса с мембраной уменьшается. Можно предположить, что участки связывания фермента на мембране несут отрицательный заряд.
Эти данные коррелируют с результатами исследования
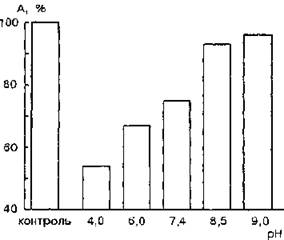
Рис. 49. Зависимость ферментативной активности лактатдегидрогеназы, связанной с мембранами эритроцитов (2-Ю-8 моль/л, 0,15 мг/мл белка мембран), от величины pH (за 100 % принята активность свободного фермента при тех же значениях pH)
В. И, Лущака (1991, 1992), который изучил влияние pH среды на экстракцию ЛДГ из микросом плавательных мышц шиповатого ската и показал, что степень экстрагируемости фермента из микросомальных мембран увеличивается при щелочных значениях pH. Т. В. Всаковаи М. В. Иванов (1992) считают, что для взаимодействия ЛДГ из скелетных мышц свиньи с мембранами саркоплазматического ретикулума важен отрицательный заряд мембраны. Кроме того, на поверхности мембраны существует положительный заряд, затрудняющий связывание ЛДГ.
Падение активности фермента в связанном состоянии может быть обусловлено следующими причинами (Б. И. Курганов, Н. И. Лобода, 1977; Б. И. Курганов, 1984):
а) стерическим экранированием активных центров при адсорбции;
б) изменением конформационного состояния белковой молекулы (или преимущественной адсорбцией одной из конформационных форм фермента, находящегося в растворе);
в) изменением микроокружения фермента при переходе из раствора в примембранный слой (концентрации водородных ионов и субстратов в растворе и на поверхности могут различаться вследствие их взаимодействия с подложкой).
Однако убедительных доказательств в пользу какого-либо одного из этих предположений не получено.
Таким образом, процесс ассоциации ЛДГ с эритроцитарной мембраной можно регулировать путем варьирования значений pH среды. Следовательно, важную роль в образовании комплексов фермента с различными клеточными структурами играют электростатические взаимодействия.
При взаимодействии ЛДГ с тенями эритроцитов, обработанными тритоном Х-100, наблюдалось снижение каталитической активности ЛДГ, однако степень инактивации фермента была выше, чем в случае его ассоциации с препаратами нативных мембран. Так, связывание ЛДГ с эритроцитарными мембранами и их “три- тоновыми” тенями при pH 7,4 приводило к снижению ее активности на 25 и 82 % соответственно (рис, 50). При pH 9,0 каталитическая активность ЛДГ в присутствии нативных и тритоновых теней статистически достоверно не отличалась от таковой для свободного фермента. Полученные нами результаты свидетельствуют о том, что ЛДГ эффективнее взаимодействует со спектрин-актино-
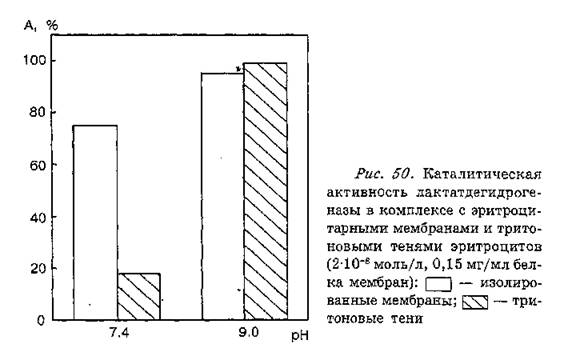
вой сетью, представляющей основу мембранного каркаса клетки. Можно предположить, что преобладающий вклад в образование комплекса фермента с эритроцитарной мембраной вносит его ассоциация с элементами цитоскелета, прежде всего актином и спектри- ном. Это соответствует концепции о центральной роли актина как белка-координатора в регуляции обменных процессов в клетке, предложенной Б. Ф. Поглазовым и соавт. (1983).
При взаимодействии эритроцитарной ЛДГ с нативными и модифицированными тритоном Х-100 мембранами эритроцитов активность связанного фермента снижается при pH 7,4 на 21 и 30 %, а при pH 5,4 — на 35 и 41 % соответственно. Следовательно, изменения каталитической активности фермента из скелетных мышц и эритроцитов в ассоциированном с эритроцитарными мембранами состоянии имеют идентичный характер и указывают на единый механизм ассоциации мембран эритроцитов с ЛДГ из названных источников.
В присутствии восстановленного кофермента в среде инкубирования ЛДГ и мембран ее активность статистически достоверно не отличается от таковой для свободного фермента. Итак, полученные результаты можно объяснить тем, что NADH, соединяясь с апоферментом, вызывает его конформационные изменения, которые затрудняют адсорбцию- фермента на подложке биологической природы. Эти данные согласуются с результатами ряда работ
(R. L. Melnick, H. 0. Hultin, 1973; Г. Л. Ермаков, 1993; A. Dabrowska et al., 1989), в которых было показано, что NADH способствует десорбции фермента, связанного с мышечными волокнами лосося, липосомами и мембранами саркоплазматического ретикулума.
Ингибирование ЛДГ, ассоциированной с субклеточными частицами, происходит за счет увеличения константы Михаэлиса для пирувата, а не изменения максимальной скорости реакции (A. Dabrowska et al., 1989). Н. П. Сугробовой и соавт. (1983) показано, что связывание ЛДГ с F-актином не изменяет характера зависимости скорости реакции от концентрации субстрата — а-кето- глутарата. В результате взаимодействия ЛДГ с F-актином максимальная скорость реакции снижается в 1,9 раза, а константа Михаэлиса увеличивается в 2,5 раза. Другие результаты получены при исследовании ЛДГ, ассоциированной с митохондриальной фракцией печени цыпленка (М. L. Sagrista, J. Bosal, 1987). Зависимость каталитической активности связанного фермента от концентрации пирувата отклоняется от гиперболической формы, а при изменении концентрации NADH описывается в рамках кинетики Михаэлиса—Ментен.
Считают, что имеющиеся в литературе противоречивые данные по этому вопросу можно объяснить двумя причинами: особенностями природы биологической подложки и природой самого фермента.
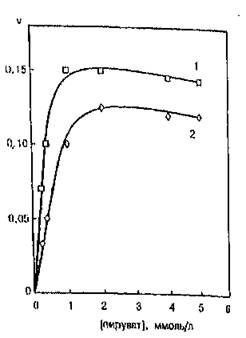
На рис. 51 показаны результаты исследования зависимости скорости ферментативной реакции ЛДГ, связанной с мембранами эритроцитов, от концентрации пирувата натрия при pH 7,4. Кинетические кривые для свободного фермента и комплекса фермент—мембрана подчиняются уравнению Михаэлиса—Ментен. Величина константы Михаэлиса для ЛДГ в растворе равна 0,25±0,02 ммоль/л пирувата, а для мембраносвязанной — 0,60±0,04 ммоль/л. Максимальная скорость ферментативной реакции при взаимодействии ЛДГ с мембраной эритроцитов снижается в 1,2 раза. Значит, ингибирование фермента происходит за счет увеличения в 2,4 раза константы Михаэлиса для пирувата натрия.
Суммируя результаты проведенных экспериментов, можно констатировать, что ассоциация ЛДГ с эритроцитарными мембранами и их белковыми компонентами, представляющими собой мембранный каркас клетки, при различных значениях pH среды приводит к снижению каталитической активности фермента, ко-
Рис, 51. Зависимость скорости ферментативной реакции лактитдогидрогвназы в свободном состоянии (!) и в комплексе с эритроцитарными мембранами (2) от концентрации пирувата натрия
торос происходит за спет увеличения константы Михаэлиса для пирувата натрия. Эффект ингибирования ЛДГ усиливается при сдвиге pH в кислую область, что свидетельствует об электростатической природе взаимодействий между ферментом и мембраной. Следовательно, процесс образования комплекса ЛДГ с биологической подложкой можно регулировать путем варьирования концентрации водородных ионов, а также добавления в реакционную среду кофермента NADH.
Обратимое взаимодействие белков-ферментов с различными структурными компонентами клетки представляет собой один из эффективных механизмов регуляции каталитической активности компонентов метаболических систем. В соответствии с отим функционирование ферментов в клетке можно рассматривать как совокупность взаимосвязанных процессов перехода их из свободного состояния в связанное, которые регулируются путем воздействия различных физико-химических факторов и сопровождаются изменением функциональных свойств этих ферментов, а, следовательно, и изменением жизнедеятельности целой клетки.
Па рис. 52, и представлены результаты исследования каталитической активности ЛДГ, УФ-облученной в дозе 1,5 кДж/и‘ и ассоциированной с эритроцитарными мембранами при pH 7,4. Активность фермента в свободном состоянии после его облучения составляет НЗ % по отношению к контрольному образцу.
1 ко
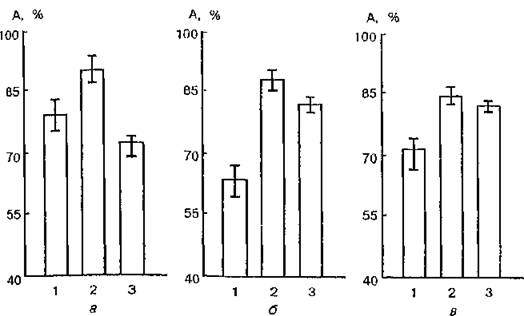
Рис, 52. Активность лактатдегидрогеназы, УФ-облучеиной в дозе 1,5 кДзк/м2, в комплексе с эритроцитарными мембранами при pH 7,4 (а), 5,4 (б), 4,5 (в): 1 — активность УФ-облученной ЛДГ; 2 — активность ЛДГ в присутствии эритроцитарных мембран; 3 — активность ЛДГ, УФ-облучспной в дозе 1,5 кДзк/м2 и ассоциированной с эритроцитарными мембранами
Каталитическая активность УФ-модифицированной ЛДГ, ассоциированной с эритроцитарными мембранами, статистически достоверно уменьшается па 8 % по сравнению с активностью свободного УФ-облученного фермента.
Таким образом, степень инактивации УФ-модифицированной ЛДГ при pH 7,4 в связанном с эритроцитарными мембранами состоянии выше, чем в свободном состоянии (табл. 13).
На рис. 52, б приведены результаты исследования каталитической активности ЛДГ, модифицированной УФ-излучением и ассоциированной с эритроцитарными мембранами при pH 5,4. Активность фермента в свободном состоянии после его УФ-облучения составляет 65 % по отношению к контрольному образцу. Каталитическая активи ость УФ-облученной ЛДГ и ассоциированной с эритроцитарными мембранами увеличивается на 21 % по сравнению с каталитической активностью УФ-облученного свободного фермента.
Другими словами, степень инактивации ЛДГ, УФ-облученной и связанной с эритроцитарными мембранами, при pH 5,4 ниже, чем при облучении свободного фермента (см. табл. 13).
Таблица 13
Степень инактивации ЛДГ, УФ-облученной при pH 7,4; 5,4; 4,5 в свободном состоянии и в комплексе с эритроцитарными мембранами
| pH | Степень инактивации ДА = Анат- А^, % | |
| в свободном состоянии | в связанном с эритроцитарными мембранами состоянии | |
| 7,4 | 17 | 21 |
| 5,4 | 35 | 6 |
| 4,5 | 14 | 2 |
Примечание. Анаг — каталитическая активность ЛДГ в нативном состоянии; А — каталитическая активность УФ-облученной ЛДГ; ДА — разность величин каталитической активности ЛДГ в нативном и УФ-облученном состояниях.
Нарис. 52, в показаны результаты исследования каталитической активности ЛДГ, УФ-облученной и ассоциированной с эритроцитарными мембранами при pH 4,5. Каталитическая активность УФ-облученной ЛДГ в комплексе с эритроцитарными мембранами при этом значении pH статистически достоверно увеличивается на 12 % по сравнению с активностью свободного УФ-облученного фермента. Степень инактивации ЛДГ, модифицированной УФ-светом и связанной с эритроцитарными мембранами, оказывается ниже, чем при облучении свободного фермента (см. табл. 13).
Следовательно, степень инактивации ЛДГ, УФ-облученной и ассоциированной с эритроцитарными мембранами, при pH 4,5 и 5,4 уменьшается по сравнению с таковой для УФ-модифициро- ванного свободного фермента при тех же значениях pH.
Можно предположить, что такое изменение функциональной активности ЛДГ, модифицированной воздействием УФ-света и ассоциированной с эритроцитарными мембранами, может быть связано с изменением микроокружения активного центра УФ- облученного фермента при его адсорбции на мембране.
Вероятно, УФ-модификация и ассоциация ЛДГ с эритроцитарной мембраной при pH 7,4 приводит к стерическому экранированию активных центров ЛДГ, а при pH 5,4 и 4,5 создаются более оптимальные условия для катализа. Но нельзя исключить и то обстоятельство, что при УФ-облучении и последующей иммобилизации фермента на мембране происходит изменение участков его локализации на подложке биологической природы.
С целью проверки справедливости высказанных предположений были исследованы флуоресцентные свойства 1-анилинона- фталин-8-сульфоната в комплексе с мембранами эритроцитов и ЛДГ, УФ-облученной при значениях pH 7,4; 5,4 и 4,5.
На рис. 53 представлены результаты исследования значений интенсивности в максимуме флуоресценции АНС (479 нм) в присутствии мембран эритроцитов и ЛДГ, модифицированной воздействием УФ-облучения при pH 7,4; 5,4 и 4,5.
Из анализа рисунка следует, что при взаимодействии эритроцитарных мембран с ЛДГ, облученной УФ-светом при указанных значениях pH, не выявляются статистически достоверные отличия значений интенсивности флуоресценции зонда в комплексе “мембраны — ЛДГ в нативном состоянии” и в комплексе “мембраны — УФ-облученный фермент”.
Полученные данные, вероятно, указывают на то, что при УФ- облучении фермента не происходит модификация тех его участков, которые ответственны за связывание с эритроцитарными мембранами.
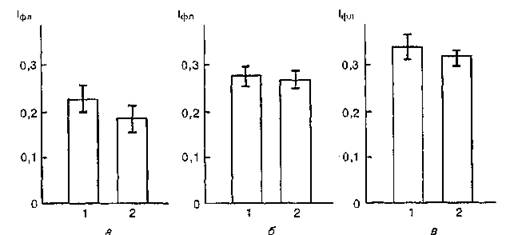
Рис. S3. Интенсивность флуоресценции зонда 1,8-АНС в комплексе с эритроцитарными мембранами и ЛДГ, модифицированной УФ-излучением, при pH 7,4 (й), 5,4 (