Структура, функциональные и некоторые физикохимические свойства Na+, К[1] АТФазы
В 1957 г. J. Skou* обнаружил в гомогенате периферических нервов краба АТФазу, активируемую ионами Na+ и К+ и ингибируемую специфическим блокатором активного транспорта одно-
Таблица 5
Некоторые ЛТФазы, участвующие в транспорте ионов
| Тип АТФаз и транспортируемые ионы | Источник | Мембрана |
| 1. Е/Е^тип | ||
| Н+ | Низшие эукариоты дрожжи и грибы | Плазматическая |
| Н+ | Высшие эукариоты Растения Животные | Плазматическая |
| к+ | Прокариоты Е. coli Высшие эукариоты животные | Ци т оп лазмати - ческая Плазматическая |
| Na+/K+ | « | « |
| Са2+ | « | « |
| Са2+ | « | Сарк оплазмати- ческая |
| 2. FjFg-тип | ||
| Н+ | Большинство бактерий | Цитоплазматиче ская |
| Н+ | Эукариоты животные и растения | Внутренняя ми- тохон дри ал ьн ая |
| н+ | Растения | Тилакоиды хлоропластов |
| 3. Вакуолярный тип | ||
| Н+ | Низшие эукариоты дрожжи и грибы | Вакуоли |
| н+ | Высшие эукариоты растения | Тонопласты |
| н+ | Высшие эукариоты животные | Лизосомы |
| н+ | « | Секреторные гранулы |
| н+ | « | Запасающие гранулы |
| н+ | « | Окаймленные везикулы |
валентных катионов — уабаином.
В 70-е гг. L. Е. Hokin было показано, что Na+, К+-АТФаза, очищенная до гомогенного состояния, при встраивании в лецитиновые липосомы способна обеспечить сопряжение гидролиза АТР с активным переносом (навстречу друг ДРУГУ) ионов Na+ и К+ со стехиометрией Na+/K+/ATP = 3/2/1, т.е. той же, что обнаружена и для нативных мембран (эритроцитов). Основной функцией Na+, К+-АТФазы (Na4-насоса), присутствующей в мембранах большинства эукариотических клеток, является установление градиентов ионов Na+ и К+, что служит необходимым условием регуляции объема клетки, внутриклеточного значения pH, процессов дыхания и гликолиза, трансмембранного переноса сахаров, аминокислот и нейротрансмиттеров. В возбудимых клетках градиент Na+ составляет движущую силу для быстрого его тока внутрь клетки при возбуждении (потенциал действия), для контроля за внутриклеточным содержанием Са2+ через Na'/Ca2+-обменный механизм.Na+, К+-АТФаза локализована в различных органах и тканях. Особенно велико ее содержание в органах, осуществляющих выделительную функцию (почки, солевые железы) или выполняющих электрическую работу (мозг, электрические органы).
Na+, К+-АТФаза всегда обнаруживается в наружных плазматических мембранах клеток и является их маркерным ферментом. Она представляет собой интегральный белок, пересекающий мембрану насквозь, контактируя как с внеклеточной средой, так и с цитоплазмой. Гидролитический центр фермента “обращен” внутрь клетки. Изнутри осуществляется также и активация фермента натрием. Присутствие калия требуется снаружи. Как гидролитический центр, так и участки ионной активации располагаются в гидрофильном окружении в тех частях молекулы, которые выступают из фосфолипидного бислоя (рис. 8).
Молекула фермента состоит из двух субъединиц: а (липопротеидной природы), содержащей гидролитический центр, способный фосфорилироваться в присутствии АТР, и р (гликопротеин), а-субъединица (каталитическая) “прошивает” липидный матрикс и несет на себе каталитический центр, центры связывания и переноса ионов натрия и калия, уабаина и других сердечных гликозидов (дигиталис-рецептор), ионов магния, других функционально важных продуктов, р-субъединица обеспечивает правильную ориентацию Na+, К+-АТФазы, отвечает за ее антигенные свойства, участвует в регуляции связывания ионов калия с ферментом.
Сиалогликопротеидный компонент обнаружен и в а-субъеди- нице Na+, К+-АТФазы почек кролика, электрического органа рыб и наплиуса креветок. Считают, что а-субъединица, судя по аминокислотному составу, более гидрофобна, хотя р-субъединица связывает большее количество фосфолипидов (табл. 6). Методом амперометрического титрования нитратом серебра установлено наличие в Na+, К+-АТФазе почек свиньи пяти дисульфидных свя-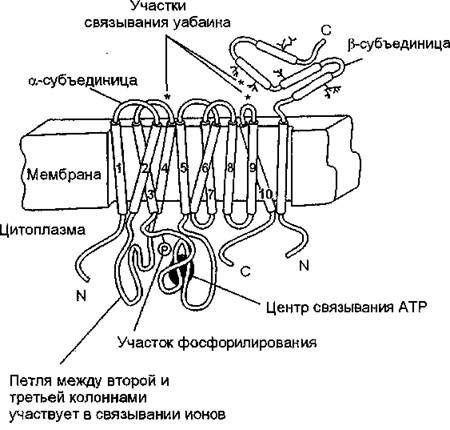
Рис. 8. Схема расположения Na+, К+-АТФазы в клеточной мембране (А. А. Болдырев, 1998)
зей и 20 свободных остатков цистеина, большинство из которых относится к разряду замаскированных.
Анализ профиля гидрофобности показывает, что в полипеп- тйдной цепи ос-субъединицы содержится от 6 до 10 потенциальных трансмембранных фрагментов (колонн), состоящих из 17 — 25 аминокислот, уложенных в сс-спираль (см. рис. 8). N- и С-кон- цевые фрагменты ос-субъединицы располагаются в цитоплазме. N- конц'евая часть полипептидной цепи представляет собой гибкий неспирализованный участок, обогащенный остатками лизина, который принимает участие в конформационных переходах и, возможно, регулирует чувствительность фермента к катионам. В N-концевой половине ос-субъединицы присутствует 4 трансмембранных фрагмента (Ml—М4), а в С-концевой половине, по-видимому, еще 6 (М4—М10). Между трансмембранными фрагментами М2 и М3 располагается малая цитоплазматическая петля, а между М4 и М5 — большой цитоплазматический домен, в состав которо-
Таблица 6
Фосфолипидный и углеводный состав изолированных а- и ft-субъединиц Na'~, К+АТФазы (А. А. Болдырев, 1985)
| Фосфолипиды, % | а-Субъединица | р-Субъединица |
| Сфингомиелин | 23 ± б | 17 ± 4 |
| Фосфатидилхолин | 31 ± 5 | 37 ± 2 |
| Фосфатидилсерин | 11 ± 2 | 11 ± 2,5 |
| Фосфатидилинозитол | 1 ± 1 | 1 ± 1 |
| Фосфатидилэтаноламин | 33 ± 7 | 33 ±4,5 |
| Углеводы, моль/100 молей аминокислот | а-субъединица | р-субъединица |
| Манноза | 0,3 ± 0,1 | 2,5 ±0,1 |
| Галактоза | 0,9 ±0,1 | 5,5 ± 0,4 |
| Глюкоза | 0,9 ±0,2 | 2,0 ± 0,5 |
| Глюкозамин | 2,0 ± 0,2 | 10,1 ± 0,5 |
| Галактозамин | Отсутствует | 0,3 ± 0,1 |
| Сиаловые кислоты | 0,35 ±0,05 | 3,2 ± 0,2 |
го входит более 400 аминокислотных остатков а-субъединицы.
Большой цитоплазматический домен состоит из чередующихся а- спиральных и Р-складчатых участков, Характерной его особенностью является наличие центрального ядра, представленного (3-структурой, которое окружено а-спиральными участками, соединенными гибкими петлями. АТР-связывающий центр расположен, вероятно, в С-концевой части (3-структуры. В большом цитоплазматическом домене ос-субъединицы находится фосфорилируемый остаток аспарагиновой кислоты (Asp 369). Полипептидная цепь Р-субъединицы уложена в антипараллельные p-структуры. Все изоформы Р-субъединицы содержат три дисульфидные связи (Cysl21— Cysl50, Cysl60—Cysl76, Cys215—Cys278 в рі-изоформє).Молекулярная масса белковых субъединиц в среднем составляет 90—130 (ос) и 35—57 (р) кДа. В очищенных препаратах Na+, К+-АТФазы рядом исследователей отмечено присутствие низкомолекулярного протеолипида с молекулярной массой 10000—15000 выраженной гидрофобной природы. Его называют у-субъединицей Na+-Hacoca. Предполагают, что подобные протеолипиды способствуют образованию трансмембранных ионных каналов в мембране или обеспечивают взаимодействие олигомерных белков с бислоем.
Основной формой Na+, К+-АТФазы, встречающейся у млекопитающих, является изофермент аїрі-типа. В 1986 г. G. Е. Shull et al. опубликовали данные о полной последовательности трех клонов кДНК из мозга крысы, соответствующих трем различным изоформам каталитической субъединицы белка (al, ос2, аЗ) и представляющих собой продукты трех неодинаковых генов. У различных видов животных обнаружено пять изоформ р-субъ- единицы, входящих в состав Na+, К+-АТФазы и Н+, К+-АТФазы ({11,(32, рЗ,Н, КрирЫ).
Деление протомера фермента на а- и [1-субъединицы условно: под электронным микроскопом оф-протомер выглядит как цилиндр с диаметром 5,4 нм и высотой 8,0 нм. Для проявления функциональной активности фермента необходим по меньшей мере димер с молекулярной массой 265 ООО.
Однако в последнее время получены активные препараты Na+, К+-АТФазы, находящиеся в мономерном состоянии.Na+, К+-АТФаза представляет собой векторную систему первично-активного транспорта, обеспечивающую сопряжение энергии ферментативного гидролиза АТР с трансмембранным проти- воградиентным пеценосом Na+ и К+:
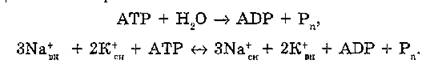
Свободная энергия гидролиза АТР составляет в условиях клетки ~ 55 кДж/моль, а для транспорта указанных ионов требуется - 46 кДж/моль, т.е. коэффициент полезного действия натриевого насоса равен 84 %. Натриевый насос работает в электроген- ном режиме: за каждый транспортный цикл из клетки “выносится” один положительный заряд. В результате на внутренней стороне плазматической мембраны создается отрицательный потенциал порядка 90 мВ.
Используя технику миллипорового фильтрования, Y. Топо- mura (1979) обнаружил в ферменте, выделенном в чистом виде из почек свиньи, участки специфического связывания Na+ (3 мо- ля/моль фермента) и К+ (2 моля/моль фермента), что согласуется как со стехиометрией транспорта ионов в оптимальных условиях (Na+/K+/ATP = 3/2/1), так и с результатами определения коэффициентов кооперативности для ионов, рассчитываемых по активации калием и натрием АТФазной реакции. Коэффициенты Хилла по ионам равны: пи по К+ - 1,7; по Na+ - 2,3. К, для Na+ в Na'-центрах составляет 0,2—0,3 ммоль/л, а в К^-центре — 2,2 ммоль/л. Ион К+ обнаруживает высокое сродство не только к собственным центрам (Kd = 0,04 ммоль/л), но и к Na'-центрам. Каждый протомер Na1, К+-АТФазы содержит один нуклеотид- связывающий центр, локализованный на а-субъединице. В присутствии Na', Mg2' , ATP он может фосфорилироваться, образуя ацилфосфат на карбоксиле аспарагиновой кислоты.
Образование фосфофермента — одна из стадий гидролиза субстрата Na', К+-АТФазой, а К1 обеспечивает его быстрое дефосфорилирование (Kd для АТФ — 0,1-4.,0 мкмоль/л). Показано существование двух типов ATP-связывающих центров, что позволяет получить двухфазную кривую субстратной зависимости фермента.Предложены три гипотезы функционирования фермента (R. W. Albers — R. L. Post, 1969; J. D. Robinson, 1975; К. R. H. Rep- ke, 1979), основанные на результатах как экспериментальных исследований, так и теоретического анализа.
В основу реакционной схемы (A. G. Sen, R. W. Albers, R. L. Post, 1969), включающей стадию, протекание которой требует наличия высокой концентрации Mg21 , положен тот факт, что для процесса фосфорилирования требуется присутствие гораздо меньшего количества Mg21 , чем для полного гидролитического цикла: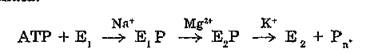
Однако этой гипотезе противоречили некоторые экспериментальные данные: одновременная, а не последовательная активация АТФазы ионами Na+ и К1 (J. D, Robinson, 1983); образование в реакционном цикле фосфофермента Et Р до фосфофермента Е2Р в отсутствие К1 (Y. Tonomura, 1973).
В соответствии со схемой J. D. Robinson (1975)
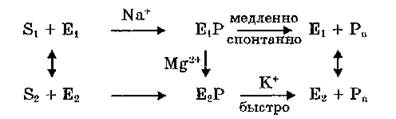
существуют две группы субстратов, гидролизующихся ферментом разными путями: “калиевые субстраты” (S2) должны взаимодействовать с конформацией фермента Е2, “натриевые субстраты” — с конформацией Е,. АТР проходит путь, включающий обе конформации фермента и стадии активации обоими ионами. Переход Е, Е, сопряжен с отщеплением ADP и изменением сродства к Na+ и К+.
Эта схема согласуется с представлениями о функционировании Na+, К+-АТФазы в виде димера, протомеры которого работают согласованно, но находятся на разных стадиях ферментативного цикла: активация натрием одного протомера совпадает по времени с активацией калием другого.
Согласно гипотезе К. R. Н. Repke (1979) Na+, К+-АТФазная активность осуществляется мономерной формой фермента. Присутствие К+ переводит фермент в димерную форму. В этих условиях присоединение АТР к центру с низким сродством к субстрату индуцирует “сбрасывание” ADP с соседнего центра, обладающего высоким сродством. Это синхронизирует работу димера в противофазе так, что эндергоническая стадия ферментативной реакции у одного протомера совпадает по времени с экзергонической стадией реакции, осуществляемой другим протомером. В пользу этой гипотезы служат результаты исследований, свидетельствующие о существовании геометрических различий между нуклеотидсвязывающими центрами двух типов, соответствующих двум значениям Км для АТР,
Однако различные модификации схемы R. W. Albers — R. L. Post не имеют принципиальной разницы и основаны на воззрениях, согласно которым каталитический цикл Na+, К+-АТФа- зы представляет собой чередование двух основных конформаций фермента (Ej и Е2) в фосфорилированном и дефосфорилирован- ном состояниях:
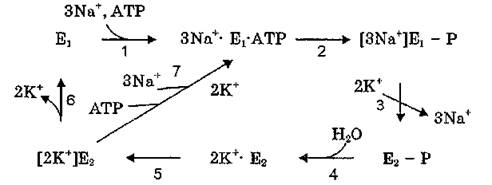
Катализ начинается, когда фермент находится в конформации Ег Эта конформация обладает высоким сродством к АТР и
ионам натрия с цитоплазматической стороны мембраны. Связывание ионов натрия активирует фосфорилирование фермента по остатку аспарагиновой кислоты. Добавление ионов калия к фос- фоферменту активирует гидролиз ацилфосфатной связи и приводит к освобождению неорганического фосфата, причем ион К' действует с внеклеточной стороны мембраны. В формировании представлений о механизме переноса ионов через мембрану сыграло свою роль открытие в середине 80-х гг. окклюдированных (поглощенных) катионов. Было установлено, что на определенных стадиях каталитического цикла ионы натрия и калия оказываются недоступными с обеих сторон мембраны, т.е, окклюдированы внутри последней.
Открытие окклюдированных ионов явилось дополнительным аргументом в пользу модели транспорта катионов Na+, К+-АТФа- зой, описывающей перенос катионов через мембрану как их перемещение через канал с калиткой (Р. Lauger, 1991). Согласно этой модели (рис. 9) катионы перемещаются через канал, образуемый полипептидной цепью а-субъединицы фермента. При этом вокруг катиона происходит формирование энергетического барьера (калитки), которая открывается и закрывается при фосфорилировании — дефосфорилировании белка. Собственно перемещение ионов представляет собой серию дискретных стадий, когда катион, соединившийся с катион-связывающим центром (при этом канал открыт только с одной стороны), переходит в окклюдированное состояние внутри канала (канал открыт .с обеих сторон), а затем освобождается с противоположной стороны мембраны (канал открыт с противоположной стороны мембраны).
Т. Нехшп et al. (1970), изучая влияние pH на Кк и vmax мембраносвязанной Na4, К'1-АТФазы из ткани мозга крысы, показали наличие в фермент-субстратном комплексе ионизирующихся групп со значениями рК: 7,1; 7,5; 7,6, которые соответствуют величинам рК для аминогрупп гистидина, цистина и «-аминокислот соответственно.
А. В. Кравцовым (1978) при исследовании солюбилизированной дигитонином Na+, К+-АТФазы из ткани мозга быка было показано, что в фермент-субстратном комплексе выявляются две ионизирующиеся группы со значениями рК 7,25 и 7,70 (т.е. близкими к приведенным выше значениям для мембраносвязанного фермента) — рис. 10, а. Величины рК этих групп соответствуют значениям для a-NH2-rpynn аминокислот и а-ЫН2-группы цис-
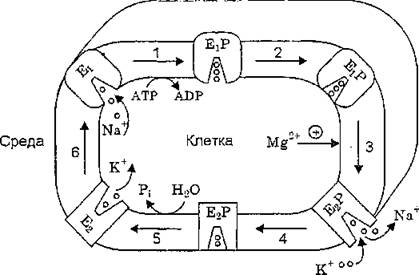
Рис. 9. Схема реакционного цикла Na+, K+-АТФазы (А. А. Болдырев, 1998)
Примечание. Шесть основных последовательных реакций включают связывание ионов натрия Е^конформером, его взаимодействие с АТР и образование фосфорилированного интермедиата (стадия 1), окклюзию ионов Na+ конформацией EtP (стадия 2), активируемый ионами Mg2+ переход BtP -> В2Р, приводящий к высвобождению ионов Na+ во внеклеточную среду и связыванию с ионным центром внеклеточного калия (стадия 3), окклюдирование ионов К+ (стадия 4), дефосфорилирование фермента, вследствие которого ионы К+ высвобождаются во внутриклеточное пространство (стадия 5), и переход конформации В2 в конформацию Ер обусловливающий начало нового цикла (стадия 6).
тина. КМАТр для солюбилизированной Na+, К+-АТФазы зависит от pH среды: при переходе от физиологических значений pH в щелочную область Км существенно уменьшается (рис. 10, б). Кривая зависимости рКм от величины pH представляет собой восходящую ветвь, что свидетельствует о присутствии в фермент-суб- стратном комплексе ионизирующейся группы с рК 7,60. Таким образом, и в случае гидролиза субстрата солюбилизированным ферментом процесс контролируется аминогруппами со значениями рК в узком интервале pH: 7,25 — 7,70.
К моменту создания модели активного центра Na1, К'-АТФа- зы, реконструированной с помощью методов ЯМР- и ЭПР-спект- роскопии, не была установлена природа группы, высвобождающей протон при переходе из К-формы в Na-форму (или акцептирующей его при превращении). Ею может быть NH2-rpynna ли-
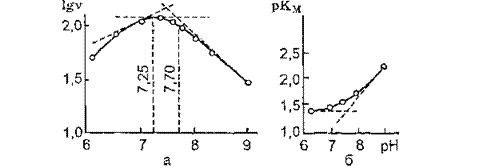
Рис. 10. Зависимость активности солюбилизированной Na+, К’-АТФя- зы от pH: а - pH-зависимость Na1, КААТФазы (lg v - f(pH)); б - зависимость KJATP) Na+, К+-АТФазы от pH
зина (J. Skou, 1983) или ОН-группа тирозина, имеющая близкое значение рК (-8,0). Обе группы идентифицированы в активном центре Na+, К?-АТФазы.
Для Na1, К'-АТФазы обнаружена нелинейная зависимость ферментативной активности от температуры в координатах Аррениуса (рис. 11). В температурном интервале 10 — 35 "С наблюдается “перегиб” на графике Аррениуса в области 20 °С, при этом энергия активации каталитической реакции повышается с 18 ккал/моль при температурах выше 20 °С до 30 ккал/моль при более низких температурах (А. А. Болдырев, 1988).
тельного изучения температурной зависимости активности Na1', Ю-АТФазы и исследования методом ЭПР параметров, характеризующих структурное состояние мембранных липидов, автором сделано заключение о том, что конформационный переход молекул фермента при изменении температуры в состояние с более высокой энергией активации обусловлен фазовыми перестройками липидного окружения при условии взаимодействия белковых молекул с липидным матриксом с участием гидрофобных и электростатических сил. Примечательным
На основании результатов сравни-
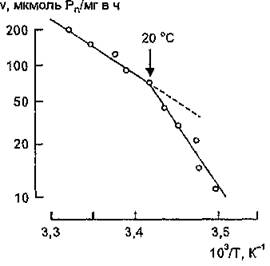
Рис. 11. Температурная зависимость начальной скорости гидролиза АТР Na+, К+-АТФазой мозга
является тот факт, что нелинейный характер графиков Аррениуса для Na+, К+-АТФазы наблюдается лишь в случае использования в качестве субстрата АТР, что свидетельствует о выполнении последним не только функции субстрата, но и модификатора.
Na+, К+-АТФаза способна гидролизовать большое количество субстратов: нуклеозидтрифосфаты, ацетилфосфат, умбеллиферон- фосфат, карбамоилфосфат, динитрофенилфосфат, (3-фурилакри- лоилфосфат. Сложная кинетика ферментативной реакции, характеризующаяся наличием двух значений Км, проявляется только при гидролизе АТР и СТР; превращения других субстратов подчиняются кинетике Михаэлиса. Na+, К+-АТФаза активируется при одновременном присутствии ионов Na+ и К? в среде. Вид зависимости активности фермента от соотношения Na+/K+ определяется природой субстрата реакции.
Считают, что структурной единицей Na+, К+-АТФазы является димер (оф)2, а минимальной функциональной единицей — (оф)-протомер (или сх-субъединица). Олигомерный ансамбль ((оср)2-димер) АТФазы поддерживается в основном за счет взаимодействий а-субъединиц с цитоплазматической стороны в “районе” ATP-связывающих центров, что обеспечивает стабильность четвертичной структуры, необходимой для проявления функциональной активности белка. Вместе с тем с функциональной точки зрения каждая а-субъединица в стабилизированном состоянии обладает полной гидролитической и транспортной активностью. Однако вопрос о биологической роли олигомеров фермента, образуемых в мембране, изучен далеко не полностью. По-видимому, наличие олигомерной структуры Na+, К+-АТФазы обеспечивает возможность реализации “гибких механизмов”, контролирующих активность мембраносвязанного фермента (см. раздел 2.3.3).
Данные, касающиеся кооперативных взаимодействий между ионными центрами и гидролизующими субстрат участками, служат основой для представлений о том, что Na+, К+-АТФаза не только структурно организована, но и использует взаимодействия между протомерами для регуляции их активности. При этом величины коэффициента Хилла пи по ионам-активаторам отражают взаимодействие между ионными центрами, локализованными на каждом протомере, а по субстрату — взаимодействие между протомерами. Чувствительность пн по АТР к фазовому состоянию мембранных липидов подтверждает, что эта величина отражает взаимодействие разных протомеров АТФазного комп
лекса. Напротив, независимость величин п„ по ионам-активаторам от фазового состояния мембраны свидетельствует о том, что взаимодействующие ионные центры находятся на одном и том же протомере.
Характеризуя натриевый насос эритроцитов, I. М. Glynn и S. J. Karlisli указали на существование четырех “дискретных режимов” его работы, соответствующих определенным частным биохимическим реакциям АТР-фосфогидролазной последовательности: 1) Na1 /К1 -обмена; 2) несопряженного выхода из клетки Na+; 3) Na'/Na4-o6MeHa; 4) К+/К4-обмена.
При отклонении от оптимальных условий насосная функция Na4, К+-АТФазы существенно изменяется. В отсутствие К4 во внешней среде и в присутствии Na4 с обеих сторон мембраны система осуществляет эквимолярный обмен ионов Na4' через мембрану, измеряемый с помощью изотопов Na+. Для этого процесса требуется присутствие АТР и АНР. Негидролизуемые аналоги АТР, в том числе 3,7-NH-ATP, заменить аденозинтрифосфат не могут (I. М. Karlish, S. Y. Glynn, 1975). Na4/Na4-o6MeH сопровождается АТР /ADP-обменом: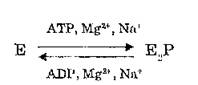
Аналогичным образом в отсутствие внутриклеточного Na1, но в присутствии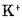 с обеих сторон мембраны Na4'-насос осуществляет неэлектрогенный обмен К1 через мембрану, не приводящий к созданию калиевого градиента. При этом требуется присутствие внутриклеточного Рп или АТР.
с обеих сторон мембраны Na4'-насос осуществляет неэлектрогенный обмен К1 через мембрану, не приводящий к созданию калиевого градиента. При этом требуется присутствие внутриклеточного Рп или АТР.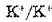 -обмен осуществляется одновре
-обмен осуществляется одновре
менно с реакцией: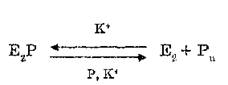
В отсутствие К? и Na+ во внешней среде Na4-nacoc может обеспечивать несопряженный выброс Na+, осуществляющийся в процессе гидролиза АТР. Стехиометрия работы насоса в этом режиме составляет 2—3 иона Na+ на 1 моль АТР.
Ш'-АТФазная реакция отличается по ряду свойств от Na+, К+- АТФазной реакции: она подчиняется кинетике Михаэлиса, имеет линейный график Аррениуса, требует меньшей концентрации MgCl2 и ингибируется высокими концентрациями АТР, хотя почти не-
чувствительна к Рп. Разница между несопряженным и сопряженным выбросами Na+ состоит в чувствительности к АТР. Несопряженный выброс Na+ достигает максимальной скорости уже при 1 мкмоль/л АТР, а Ыа+/К+-обмен требует миллимолярных концентраций субстрата. Следовательно, два типа функционирования Ыа+-насоса, как и два способа работы АТФазы, отличаются функционированием лишь субстратных центров с высоким сродством (Na", К+-АТФаза) или обоих типов субстратных центров.
Чувствительность Na+, К4-АТФазы и Na+-Hacoca к одновалентным катионам одинакова. Этот факт можно рассматривать как доказательство идентичности центров активации Na+, К+-АТФа- зы натрием и калием и центров, обеспечивающих их транслокацию. Об этом же свидетельствует соответствие стехиометрии транспорта ионов (Na+/K+ = 3/2) количеству ионсвязывающих участков на одной молекуле фермента.
Процесс переноса ионов через мембрану с помощью натриевого насоса не требует затрат энергии, если исключить работу по переносу нескомпенсированного заряда Na+ против трансмембранного потенциала при электрогенном транспорте. На это указывает существование в схеме натриевого насоса Na+/Na+ и К+/К+-обменов, для которых АТР требуется лишь как кофактор, но не источник энергии. Можно предполагать, что энергия АТР при работе Na+, К+-АТФазы затрачивается на “узнавание” ионов Na+ и К+, т.е. на связывание и сбрасывание ионов с “нужной” стороны мебраны. В норме катионы связываются с той ее стороны, где их мало, а сбрасываются туда, где их концентрация велика. Такие изменения сродства ионсвязывающих центров сопряжены с конформационными изменениями фермента, которые и являются главными энергоакцепторными стадиями реакции.
Ионы Na+ и К+ транспортируются Ыа+-насосом в частично дегидратированном состоянии. Модификаторы гидрофобных взаимодействий влияют в первую очередь на калиевую активацию, а вещества, разрушающие водородные связи, — на натриевую. Из этого можно сделать вывод, что связывание Na+ опосредовано “жесткими” структурами белка, стабилизированными водородными связями наподобие ионофорных структур. Ионы Na+ дегидрируются при переходе из водной фазы в полярную полость молекулы фермента. Ионы К+ дегидрируются при переходе в гидрофобную область молекулы. Следовательно, Na+, К+-АТФаза “различает” ионы Na+ и К+, используя различные пути их гидратации. При отсутствии внеклеточного К+ Na+/K+-Hacocbi могут осуществлять электрогенный транспорт протонов, и этот транспорт осуществляется, по-видимому, конформацией Е2 насосов.
К ингибиторам Na+, К+-АТФазы относятся олигомицин, уабаин, диметилсульфоксид, уксусный альдегид, дигитонин, тимеро- зал, этилмеркуриат. Вместе с тем необходимо отметить, что действие этих соединений на Na+, К+-АТФазную активность может быть различным.
Механизм действия модификаторов трудно поддается классификации: здесь есть агенты направленного действия (SH-pea- генты: тимерозал, этилмеркуриат, уксусный ангидрид), вещества, модифицирующие гидрофобные взаимодействия (диметилсульфоксид, олигомицин, дигитонин), специфический ингибитор Na+, К+/АТФазы уабаин ( строфантин G), препятствующий гидролизу фосфорилировэнного фермента. Гидролиз одним и тем же ферментом разных субстратов в различных условиях неодинаково чувствителен к модификаторам, что можно объяснить тем, что исследуемые “частные” реакции ферментативной активности осуществляются разными олигомерными состояниями Na+, К+-АТФазы.
Таким образом, на современном уровне знаний Na+, К+-АТФаза представляется олигомером, в котором взаимодействие между протомерами выражено сильнее, чем между глобулами белка и его липидным окружением. Количество протомеров в таком комплексе определяется, по-видимому, той конформацией белка, которую ему диктует липидное микроокружение. Сами взаимодействия между протомерами контролируются липидами. Показано, что взаимодействия между протомерами фермента в процессе гидролиза АТР не остаются постоянными: доля крупных ассоци- атов АТФазы возрастает на стадии взаимодействия ее с ионами калия, а подвижность этих ассоциатов в мембране резко увеличивается при связывании АТР.
Особый интерес представляет проблема вовлечения Na+, К+-АТФазы в общие пути передачи сигнала в клетке, а также ее фосфорилирование протеинкиназами и взаимодействие с внутриклеточными белками (см. главу 2). Установлено, что «-субъединица фермента является мишенью для сАМР-зависимой' протеинкиназы (протеинкиназы А) и фосфолипидзависимой протеинкиназы С и ионов кальция. Противоречивые данные (ингибирование, активация, отсутствие эффекта) о характере влияния фосфорилирования Na+, К'-АТФазы протеинкиназой А связывают с зависимостью “ответной реакции” белка от его конформа- ционного состояния, изменяющегося в результате взаимодействия фермента с белками цитоскелета, в частности, актином. Однако более детальные исследования по этому вопросу пока отсутствуют,
1.2.5.