Цитотоксическое действие АКМ
Защитная функция воспаления требует включения механизмов микробицидного, фунгицидного, цитотоксического действия, что осуществляется посредством выделения множества токсических соединений.
Повреждение клеток может быть индуцировано микробными токсинами или их ферментами, высвобождающимися из лейкоцитов гидролазами и катионными полипептидами, цитокинами, цитотоксическими антителами, белками системы комплемента, а также высокореакционными молекулами АКМ. Только нейтрофилы имеют в своем микробицидном арсенале более 50 токсинов [177, 1740], эффективных в отношении не только чужеродные микроорганизмов, но также и клеток собственных тканей. Если учесть, что эти токсины в очаге воспаления действуют одновременно, часто синергично друг с другом [658], то становится ясно, что выделить какой-то один механизм токсичности достаточно трудно.Исследования на клеточных культурах показывают, что на АКМ приходится от 40 до 90 % (в зависимости от штамма бактерий) микробицидного действия фагоцитирующих клеток. При этом все специализированные ферментативные системы синтеза АКМ (NADPH-оксидаза, МПО, ЭПО и NO-синтазы) участвуют в микробицидном действии фагоцитов. Генетические дефекты NADPH-оксидазы, в результате которых нарушается ее способность восстанавливать кислород, являются причиной хронического гранулематоза (встречается редко — 1 случай на 600 000 жителей); люди с данной
патологией умирают в раннем возрасте от бактериальных инфекций. Врожденные дефициты МПО встречается довольно часто (с частотой 1 : (2000—4000) [1200]); у людей с этим нарушением выявляется пониженная способность гранулоцитов убивать бактерии Candida albicans, хотя существенное возрастание заболеваемости инфекционными патологиями для них не характерно (за исключением больных диабетом), и клинически такие нарушения не проявляются. Генетические дефекты NO-синтазы не описаны, однако известно, что, в отличие от клеток животных, фагоциты человека экспрессируют очень низкую активность фермента [1737].
Несмотря на большое количество работ, показывающих участие ферментативных систем синтеза АКМ в микробицидном и цитотоксическом действии фагоцитирующих клеток, данный вопрос остается объектом дискуссий [492, 1740]. Дело в том, что активация NADPH-оксидазы приводит к синтезу О2, однако данный радикал обладает слабой реакционной и цитотоксической активностью, а в биохимических реакциях выступает как окислителем, так и восстановителем. NO-радикал также не является активным цитотоксином при прямом, неопосредованном действии на клетки; более того, во многих радикальных окислительных реакциях NO· выступает в качестве антиоксиданта [147]. Гипогалогениты, образующиеся при активации МПО и ЭПО, — эффективные окислители, цитотоксичные в системах in vitro, однако, как отмечалось выше, генетические дефициты МПО слабо проявляются на уровне целого организма, и роль данной ферментативной системы в защите от инфекций также неясна. Таким образом, можно сделать вывод, что ни одна из ферментативных систем генерации АКМ сама по себе не обладает существенным цитотоксическим действием.
Получение методами генной инженерии трансгенных экспериментальных животных, несущих мутации в генах NADPH-оксидазы или индуцибельной NO-синтазы, позволило лучше моделировать патологии человека и, соответственно, исследовать вклад каждой из данных ферментативных систем в защиту организма от патогенных бактерий и микроорганизмов. Однако уже первые работы в этом направлении выявили определенные несоответствия патологических моделей у животных (мыши, крысы) генетически обусловленным заболеваниям человека. В частности, у людей дефект
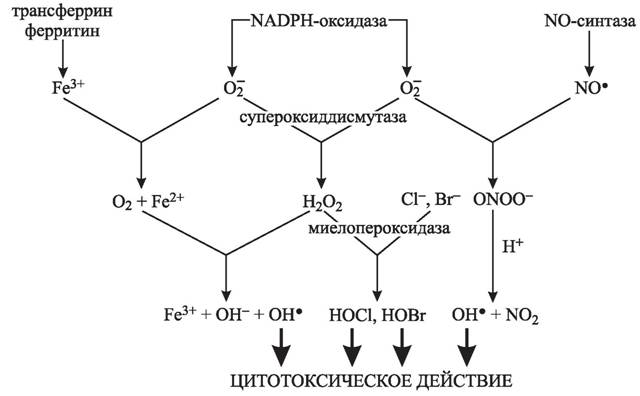
Рис. 2. Механизмы цитотоксического действия АКМ при стимуляции фагоцитов
гена CYBB, кодирующего каталитическую субъединицу gp91phox NADPH-оксидазы, сопровождается развитием хронического гранулематоза, в то время как мыши, несущие аналогичную мутацию, оказались устойчивыми к инфицированию бактериями, в том числе Salmonella typhimurium, что объясняется включением эффективных компенсаторных механизмов цитотоксического действия [1503].
Макрофаги мышей, дефицитных по NADPH-оксидазе и индуцибельной NO-синтазе, не вызывали деструкцию вирулентных бактерий Listeria, однако были достаточно эффективны в отношении бактерий S. typhimurium и E. coli, что свидетельствует о наличии в фагоцитирующих клетках других механизмов микробицидного действия, не зависящих от NADPH-оксидазы и NO-синтазы. У дефицитных по индуцибельной NO-синтазе мышей наблюдались более выраженные воспалительные повреждения, однако они были более устойчивы к развитию септического шока [700]. Для объяснения прямой зависимости микробицидного, цитотоксического и мутагенного действия фагоцитов от активности ферментативных механизмов синтеза АКМ предлагается следующая схема развития событий в процессе фагоцитоза (рис. 2).При стимуляции фагоцитирующих клеток в результате активации NADPH-оксидазы наблюдается резкое усиление продукции О2: так, 104 гранулоцитов могут синтезировать 10 нмоль радикалов в час [147] или около 2 · 108 молекул O2 в секунду на клетку. Считается, что супероксид-анион не обладает непосредственной микробицидной активностью, а является триггером каскада реакций, приводящих к образованию других более реакционных форм АКМ (ОН·, 1O2) и пероксинитрита. В присутствии СОД O2 быстро переходит в H2O2, а в реакциях с ионами металлов переменной валентности образуются синглетный кислород и гидроксильный радикал. Участие ОН-радикалов, образующихся в реакциях Габера-Вейса и Фентона, в микробицидном и туморицид- ном действии фагоцитов в настоящее время твердо доказано. Гидроксильные радикалы вызывают повреждение нуклеиновых кислот, белков и индуцируют образование органических радикалов. Оценки показывают, что цитотоксичность фагоцитов более чем на 50 % обусловлена ОН-радикалами, при этом в клетках выделяют два критических объекта повреждения: нуклеиновые кислоты и мембранные белки. Каталаза, хелаторы металлов переменной валентности, а также ингибиторы ОН-радикалов (диметилсульфоксид, диметилтиомочевина) значительно снижают цитотоксическое действие фагоцитирующих клеток, что является доказательством ключевой роли ОН-радикалов в повреждении клеток и биологических структур [901, 1721].
Взаимодействие О2 с оксидом азота приводит к образованию реакционного пероксинитрита (ONOO-). Среднее время жизни пероксинитрита в фосфатном буфере при рН 7,4 и 37 °С составляет 1-2 с, поэтому он может мигрировать в тканях. Пероксинитрит — сильный окислитель, способный окислять NH- и SH-группы белков, в результате чего инактивируется aj-ингибитор протеиназ, Mn-СОД и Fe-СОД; он индуцирует процессы ПОЛ в мембранах, вызывает однонитевые разрывы ДНК и ингибирует митохондриальное дыхание. В концентрации 250 мкМ пероксинитрит вызывал пятидесятипроцентную гибель бактерий Е. coli [1840]; в концентрациях от 1 до 100 мМ индуцировал лизис опухолевых клеток (культуры HL-60 и U-937), причем гибель клеток проходила путем классического апоптоза: с деградацией и фрагментацией ДНК, конденсацией хроматина и фрагментацией ядра [1031]. Хотя микробицидность фагоцитирующих клеток от больных с хроническим гранулематозом в отношении большинства штаммов бактерий значительно снижена, эти клетки обладают кислородзависимой токсичностью против опухолевых клеток и внутриклеточных микроорганизмов, что может быть связано с наработкой NO-радикалов.
Мишенью действия NO-радикалов являются железосерные центры ферментов, к которым принадлежат аконитаза и I, II, III комплексы цепи переноса электронов в ци- тохромоксидазе. Ингибирование окислительного фосфорилирования в митохондриях приводит к снижению внутриклеточного уровня АТФ и может являться причиной цитотоксического действия NO· как против бактерий, так и против клеток собственного
организма. Перекись водорода ингибирует глицеральдегид-3-фосфатдегидрогеназу, ключевой фермент гликолиза [292], и тем самым усиливает энергетический дисбаланс, возникающий при действии NO-радикалов. В физиологических условиях NO· взаимодействует с молекулярным кислородом с образованием оксидов азота NO2 и NO-, которые являются сильными окислителями. Цитотоксичность NO- и NO- в отношении бактерий E.
coli значительно возрастает при низких рН и в присутствии перекиси водорода и миелопероксидазы; вместе с тем нитрит снижает микробицидное действие гипохлорита в системе «Н2О2 — МПО — Cl-» [902].Дегрануляция, которой сопровождается активация гранулоцитов, приводит к высвобождению пероксидаз и инициации образования гипогалогенитов, являющихся важным компонентом микробицидного потенциала полиморфноядерных лейкоцитов [901]. Концентрация HOCl в очаге воспаления может достигать 100 мкМ [217]. Гипо- галогениты представляют собой мощные токсины, высокореакционные в химическом отношении, они взаимодействуют с мишенью либо галогенируя ее (галоген ковалентно связывается с мишенью), либо окисляя. В качестве мишеней действия системы «МПО - H2O2 - галогенид-анион» выступает чрезвычайно широкий спектр клеток и молекул. При взаимодействии с гипогалоидами в первую очередь окисляются сульфгид- рильные и тиоэфирные группы белков, поэтому наличие в среде молекул, содержащих данные группы (глутатион, альбумин, 2-нитро-5-тиобензойная кислота), существенно снижает цитотоксическое и деструктивное действие как самих гипогалоидов, так и активированных гранулоцитов. Окислению системой «МПО - H2O2 - галогенид-анион» подвержены железосерные центры и гемовые группы ферментов и металлопротеинов, при этом высвобождается железо в каталитически активной форме (Fe2+).
Несмотря на то, что цитотоксичность Н2О2 значительно возрастает в присутствии МПО и гранулоциты, которые называют клетками-«камикадзе», буквально начинены МПО (от 2 до 5 % сухого веса клетки), многие исследователи считают, что in vivo ги- погалогениты токсичны не сами по себе, а через нарушение транспортных функций мембранных белков, инактивацию АТФ-синтетазы или бактериальной убихинолок- сидазы, усиление активности протеиназ посредством инактивации их ингибиторов, активацию белков системы комплемента. Одним из возможных механизмов повреждающего действия HOCl в очаге воспаления может быть мобилизация ионов цинка из металлопротеинов, в результате чего повреждаются молекулы, содержащие связи S-Zn.
К таким молекулам относятся многие гормональные рецепторы и факторы транскрипции, металлотионеин, алкогольдегидрогеназа, коллагеназа; вместе с тем ионы цинка, входящие в состав активного центра C^Zn-СОД, устойчивы к действию HOCl, так как они не связаны с атомами серы [609]. Мобилизация цинка из белков, содержащих S-Zn связи, вызывается физиологическими концентрациями HOCl (50-200 мкМ), и в этом процессе HOCl значительно более эффективна по сравнению с О-2 и Н2О2 [609].Рассмотрение механизмов цитотоксического действия АКМ (см. рис. 2) показывает, что наибольший эффект достигается при включении нескольких механизмов образования АКМ:
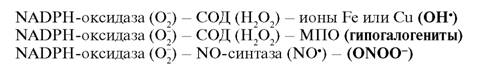
При этом можно видеть, что процесс эволюции сопровождается разнесением этих механизмов как в пространстве, так и во времени. В организме человека только в гранулоцитах находятся две эффективные ферментативные системы синтеза АКМ: NADPH- оксидаза и миелопероксидаза, поэтому для них оправдано название клетки-«камикад- зе». Моноциты имеют невысокое содержание МПО, от которой они избавляются в процессе трансформации в макрофаги. В человеческих моноцитах и макрофагах, в отличие от фагоцитов грызунов, активность индуцибельной NO-синтазы невысока [241],
вместе с тем при действии цитокинов индукция фермента выявлена в человеческих ге- патоцитах, хондроцитах, клетках поджелудочной и щитовидной желез, мезангиальных и гладкомышечных клетках, остеобластах, кардиомиоцитах. Поэтому синтез NO· этими клетками может вносить вклад в микробицидное действие в области воспаления. Если развитие дыхательного «взрыва» в фагоцитах при активации NADPH-оксидазы является быстрым процессом (как правило, от 10 минут до 2 часов), то NO-синтаза — ин- дуцибельный фермент, и ее активность возрастает после лаг-фазы (обычно несколько часов), в течение которой происходит синтез фермента.
Включение основных механизмов синтеза АКМ также находится под сложным контролем. Максимальная продукция радикалов NO-синтазой и NADPH-оксидазой наблюдается только при воздействии нескольких стимулов, например, цитокина, оказывающего примирующий эффект, и липополисахарида. Более того, NO-радикалы индуцируют синтез цитоплазматической C^Zn-СОД, что снижает образование в клетках высокотоксического пероксинитрита, в частности при эндотоксическом шоке [618]. Синтезирующийся в печени С-реактивный белок ингибирует фагоцитарную NADPH-оксидазу и индуцирует синтез NO-синтазы, что снижает повреждающее действие АКМ на гепатоциты [1381]. По-видимому, такое пространственно-временное разнесение и многоуровневая система активации необходимы для эффективного контроля над цитотоксическим действием АКМ: деструктивный эффект должен проявляться в первую очередь в отношении чужеродных микроорганизмов и в наименьшей степени затрагивать клетки и ткани собственного организма. Другим аспектом участия разных механизмов образования АКМ в цитотоксическом действии является антагонизм и синергизм действия радикалов. В частности, NO-радикал эффективно ингибирует O2 и органические радикалы, и поэтому в некоторых случаях он может рассматриваться как антиоксидант, оказывающий защитное действие [485].
В очаге воспаления фагоцитирующие клетки функционируют в условиях выраженного окислительного стресса. При активации ферментативных механизмов синтеза АКМ их токсическому действию подвержены сами фагоциты. Поэтому интересен вопрос: каким же образом фагоцитирующие клетки защищаются от АКМ? Дыхательный «взрыв» в моноцитах и макрофагах сопровождается быстрым (20—30 мин) повышением внутриклеточного содержания глутатиона, который поступает через специальные транспортеры [1489]. Прямое действие на клетки H2O2 также сопровождалось усилением транспорта глутатиона. Высказывается предположение о наличии активной защиты в условиях окислительного стресса, которая индуцируется через фактор транскрипции Nrf2 [131]. Активация фактора транскрипции NF-кВ также повышает устойчивость клеток к апоптоз-индуцирующим воздействиям, в том числе АКМ [348]. Таким образом, одновременно с усилением продукции АКМ моноциты и макрофаги повышают свой уровень антиоксидантной защиты.
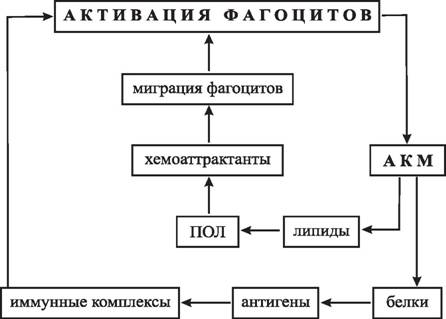
Некоторые хронические патологии могут быть следствием аутотоксического действия АКМ: так, воспаление в области поджелудочной железы часто приводит к диабету, возможной причиной этого является индукция в β-клетках NO-синтазы [938], которая совместно с NADPH-оксидазой фагоцитов вызывает деструкцию инсулин-продуцирую- щих клеток. В клетках щитовидной железы также индуцируется NO-синтаза [858], и тиреоидные гланды являются одной из главных мишеней для аутоиммунной атаки у человека. Хондроциты синтезируют NO· [1266], в результате множество людей страдают ревматоидным артритом. Кардиомиоциты, гепатоциты, гладкомышечные клетки также являются удобными объектами аутоповреждений у человека, в результате — инфаркт миокарда, цирроз печени, атеросклероз сосудов. В 1996 году выделен аутоиммунный синдром Х, для которого характерно повреждение кровеносных сосудов в результате атаки лейкоцитами крови; заболевание протекает быстро и через 3—4 дня заканчивается летальным исходом; это является ярким примером высокого цитотоксического потенциала фагоцитов. В ос-
нове развития многих аутоиммунных патологий и хронических воспалительных процессов может лежать образование так называемого «порочного круга» [41]:

выступают как поликатионные агенты и даже в денатурированной форме могут индуцировать лизис бактерий [658]. Токсичность трипсина, химотрипсина, катепсина G и панкреатической эластазы в отношении эндотелиальных клеток усиливалась низкими концентрациями Н2О2 [658]. Лизис эритроцитов активированными нейтрофилами зависел от содержания нейтральных протеиназ и активности синтеза АКМ [1338].
Эти примеры указывают на то, что выделить в чистом виде один механизм цитотоксического действия в организме нельзя, более того, на сегодняшний день наши познания еще далеки от понимания реальных ситуаций, происходящих in vivo. Так, недавно была показана возможность образования озона и озоноподобных соединений (Н2О3, НО·) в реакциях антиген-антитело в присутствии синглетного кислорода [1747]. Так как антитела (IgG) и O2 выявляются в относительно высоких концентрациях в фагосомах стимулированных нейтрофилов, то исследователи считают возможным участие озоноподобных соединений в инактивации антигенов и микробицидном действии фагоцитов [288]. Таким образом, в исследованиях механизмов деструктивного и цитотоксического действия АКМ можно ожидать появления достаточно неожиданных элементов.
Еще по теме Цитотоксическое действие АКМ:
- Микробицидное действие АКМ на микобактерии
- Защита микобактерий от токсического действия АКМ
- Почему АКМ — главное оружие фагоцитов в борьбе с микроорганизмами?
- ОКИСЛИТЕЛЬНЫЕ ПРОЦЕССЫ С УЧАСТИЕМ АКМ ПРИ ОПУХОЛЕВЫХ ПРОЦЕССАХ
- 34. Следственные действия. Виды следственных действий, участие понятых в следственных действиях. Протокол следственного действия
- Глагольное действие sub specie dverbiorum: 2. Ответные действия и языковые ответы
- 6. Как перцептивные операции и действия включаются в состав других типов человеческих действий?
- § 1. Конкретизированность, однородность и разнородность действий. Совместимость действий
- При этом судья не вправе проводить судебные действия (аналог следственных действий), направленные на исследование
- 21. Понятия и виды следственных действий. Общие условия производства следственных действий.
- Глава III. Государственные системы защиты населения и территорий страны от чрезвычайных ситуаций природного и техногенного характера и опасностей, возникающих при ведении военных действий или вследствие этих действий
- Основные правила совершения нотариальных действий. Место совершения нотариальных действий
- 3. Если семья или государство идет по неправильному пути, в семье возникают - разногласия, скандалы, драки, убийства, в государстве - кризис, войны, агрессивные действия оппозиции. Все эти действия являются показателями правильного или неправильного развития семейных, общественных отношений.
- Прежде чем приступить к рассмотрению того, как функционирует контрольный механизм Европейской конвенции о правах человека, а также тех прав и свобод, соблюдение которых государства-участники Конвенции должны гарантировать в соответствии с этим договором, следует хотя бы коротко остановиться на некоторых теоретических моментах. В частности, необходимо сказать несколько слов о структуре Конвенции, о территориальной сфере ее действия, о действии Европейской Конвенции во времени, о применении ее
- Действие.