Радиационно-химические превращения структурных компонентов биомембран
Процесс свободнорадикального пероксидного окисления липидов мембран рассматривают в настоящее время как один из механизмов “биохимического усиления” эффекта ионизирующей
радиации, который играет важную роль в развитии лучевого поражения живых систем.
При первичном взаимодействии (физическая стадия) ионизирующего излучения электромагнитной или корпускулярной природы с атомами вещества образуются положительно и отрицательно заряженные ионы, а также возбужденные электронные состояния атомов и молекул. В элементарном акте ионизации расходуется около 10—12 эВ энергии ионизирующей радиации (потенциал ионизации). Бели передаваемая электрону энергия больше этой величины, то он сам становится источником ионизации других атомов; если меньше потенциала ионизации, имеет место возбуждение атома (молекулы). Физико-химическая стадия воздействия радиации на биообъекты существенно зависит от особенностей их структурно-функциональной организации. При этом большое значение имеет процесс радиолиза воды:
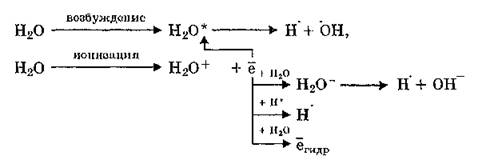
Молекулярный кислород, растворенный в биологических жидкостях, вступая в реакцию с продуктами первичного радиолиза, является источником образования супероксидного анион-радикала, гидропероксидов, органических пероксидов, эпоксидов:
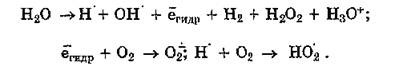
Все последующие реакции, развивающиеся в живых системах под влиянием вышеуказанных продуктов взаимодействия ионизирующей радиации с веществом, носят свободнорадикальный характер (см. главу 3). Вклад косвенного действия (т. е. с участием воды) ионизирующего излучения на биологические молекулы составляет 85—90 %. АФК, образующиеся при лучевом воздействии, индуцируют окислительные реакции в макромолекулах по оксидазному или оксигеназному пути.
В результате ферментативного или неферментативного оксигеназного окисления образуются пероксидные продукты, способные к повреждению биомакромолекул и субклеточных структур.Сравнительный анализ величин радиационно-химического выхода G (равного числу изменившихся или вновь образованных молекул на 100 эВ поглощенной энергии), используемого для характеристики радиочувствительности молекул, показал, что наиболее радиочувствительным компонентом клетки являются фосфолипиды (G>1), выступающие в качестве источника образования первичных продуктов радиолиза — органических радикалов и пероксидов, участвующих в реакциях инициации, продолжения и разветвления цепей свободнорадикального окисления.
Активные радикальные метаболиты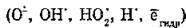 и
и
продукты радиолиза липидов мембран индуцируют в белках процессы окисления тиоловых и аминогрупп, образования внутри- и межмолекулярных сшивок, сопровождающиеся их инактивацией. В нуклеиновых кислотах и нуклеотидах происходят одно- и двутяжевые разрывы, модификация азотистых оснований, образование сшивок ДНК—ДНК и ДНК—белок.
Таким образом, в результате воздействия ионизирующей радиации на биомембраны происходит нарушение структурной организации всех ее компонентов, изменяется протекание транспортных и метаболических процессов, становится возможным выход лизосомальных ферментов (в частности, протеаз и нуклеаз), разрушение внутриклеточных структур и гибель клетки.
Важную роль в инактивации токсичных продуктов ПОЛ играют фосфолипазы, особенно фосфолипазы А2. В процессе фосфолипазного гидролиза происходит удаление из мембран наиболее глубоко деградированных фрагментов ненасыщенных жирных кислот липидов. В результате токсичные продукты ПОЛ переходят в гидрофильное окружение и подвергаются ипактивации с участием монооксигеназной системы (в том числе цитохрома Р-450) эндоплазматического ретикулума.
В 60-х гт. было показано, что ионизирующее излучение ингибирует работу натриевого насоса эритроцитов человека.
Торможение активного оттока Na+ наблюдается уже после облучения в дозах 8,9—89 Гр. При повышении дозы радиации до 200 Гр транспорт Na+ и К+ в эритроцитах полностью инактивируется. После облучения эритроцитов крысы в дозе 7 Гр также выявлено существенное торможение активности Na+, К+-АТФазы.Результаты исследования влияния общего рентгеновского облучения на функциональные свойства Na+, К+-АТФазы мембран клеток печени крысы показали, что торможение удельной, активности фермента отмечается в течение длительного времени (1ч — 60 сут) после прекращения воздействия радиации. Через 1 ч потеря активности фермента составляла 87 %. Через 30 и 60 сут у выживших животных наблюдалась полная инактивация фермента.
Следовательно, ингибирование активного мембранного транспорта под действием ионизирующего излучения происходит в клетках различных типов, в разных условиях облучения в широком диапазоне доз. Предполагают, что сохранение жизнедеятельности клеток при дезактивации натриевого насоса связано с включением компенсаторных механизмов поддержания гомеостаза. Например, в мембранах эритроцитов при торможении активности Na+, К+-АТФазы активность Са2+-АТФазы превышает контрольный уровень, а в плазматических мембранах печени увеличивается Мё2+-АТФазная активность. Известно, что Са2' и Mg2+ способствуют связыванию белков, в том числе АТФаз, с мембраной. В липидных бислоях Са2+ обеспечивает образование мостиков между фосфатидами, в результате которого упаковка липидной фазы становится более плотной и уменьшается проницаемость мембраны. Кроме того, после рентгеновского облучения животных в дозе 5 Гр обнаруживается повышение активности щелочной фосфатазы, связанной с плазматическими мембранами клеток печени мышей. Щелочная фосфатаза — интегральный фермент плазматических мембран некоторых клеток — участвует в активном транспорте ионов Na+ и К+.
Воздействие ионизирующего излучения приводит к существенному подавлению активности другого транспортного белка — НСО3"-АТФазы плазматических мембран клеток печени и эритроцитов, в результате которого возможно нарушение кислотнощелочного равновесия в клетке и организме.
Торможение функциональной активности АТФаз, участвующих в активных транспортных процессах, коррелирует с нарушением баланса электролитов в облученной клетке, а также с изменением проницаемости плазматических и внутриклеточных мембран, вследствие чего регистрируется выход внутриклеточного калия, подавление окислительного фосфорилирования, утечка К+ из митохондрий.
Функциональная активность мембраносвязанной 5'-нуклео- тидазы печени крыс значительно возрастает при действии ионизирующей радиации как в ранние, так и в поздние сроки после облучения. Считают, что повышение активности этого фермента не связано с синтезом фермента de novo, а обусловлено перераспределением белка в растворе и в составе мембран.
По-видимому, механизмы пострадиационной вариабильнос- ти активности мембранных белков предусматривают реализацию регуляторных эффектов целого ряда факторов: например, влияния нейроэндокринной системы на функционирование ферментов в облученном организме, биологически активных веществ, продуктов пероксидного окисления липидов, изменений зарядового состояния поверхностных участков мембраны и др.
Ионизирующее излучение индуцирует снижение общего уровня мембранных фосфолипидов и повышение содержания холестерина, что сопровождается возрастанием коэффициента холе- стерин/фосфолипиды до 1,05 при норме 0,60. Однако уровень индивидуальных фосфолипидов изменяется разнонаправленно: происходит накопление сфингомиелина и фосфатидилсерина и снижение содержания фосфатидилхолина, фосфатидилэтаноламина и особенно фосфатидилинозитола. В целом коэффициент насыщенности мембранных липидов повышается. Все это приводит к значительным нарушениям текучести мембраны, увеличению ее вязкости, изменению функциональных свойств мембранных белков.
Предполагают, что механизм пострадиационной модификации состава и содержания структурных липидов в плазматических мембранах животной клетки связан с изменением процессов синтеза и распада липидпереносящих белков, ферментов липидного обмена, нарушением внутримембранной динамики липидных компонентов.Вместе с тем необходимо отметить, что липиды клеточных мембран животных защищены от процесса ПОЛ при воздействии ионизирующей радиации по сравнению с искусственными мембранными структурами, в частности, липосомами. Накопление холестерина может препятствовать приросту уровня пероксидов липидов и приводить к торможению других процессов в мембране, связанных с развитием лучевой патологии. Кроме того, в мембранах локализованы собственные низкомолекулярные антиоксиданты (например, а-токоферол) и защитные антиокислительные ферментные системы. Так, результаты модельных исследований свидетельствуют о том, что витамин В образует комплексы с мембранными фосфолипидами, содержащими остатки арахидоновой кислоты. Формирование этих комплексов способствует стабилизации и снижению проницаемости клеточных мембран с высоким содержанием полиненасыщенных жирных кислот.
При изучении влияния минимальных летальных доз (154,8 мКл/кг) рентгеновского излучения на активность некоторых ферментов углеводного обмена клеток костного мозга крыс выявлено, что после облучения в популяциях миелокариоцитов активность лактат-, малат- и глюкозо-6-фосфатдегидрогеназ значительно угнетается в течение 72 ч после облучения и достигает минимального значения на 3-й сутки эксперимента. На 10-е сутки наблюдается увеличение значений скорости реакций, катализируемых. малат- и лактатдегидрогеназой, при снижении активности глюкозо-6-фосфатдегидрогеназы. Следовательно, в ранние сроки после воздействия ионизирующей радиации на организм животных происходит усиление интенсивности анаэробных процессов и ослабление дыхания и пентозофосфатного пути.
Облучение животных ионизирующей радиацией вызывает не только изменения активности многих ферментов, регулирующих процессы клеточного метаболизма, но и относительного содержания изоферментов.
Так, в ряде работ показано, что у-облучение влияет на активность изоферментов лактатдегидрогеназы, кислой фосфатазы, неспецифической эстеразы. Причем при более низких дозах облучения величина изменений в относительном содержании изоформ может быть больше, чем при высоких. При анализе относительного содержания фракций изоферментов вышеуказанных белков в различные сроки после у-облучения в дозе 1 Гр установлено, что воздействие радиации вызывает существенные изменения в относительном содержании изоформ изученных ферментов, которые имеют, по-видимому, фазовый характер. Считают, что эти результаты связаны с интенсивной перестройкой генетического аппарата клетки в период до 7 суток после облучения. Новому структурно-функциональному состоянию генетического аппарата соответствует иной уровень процессов метаболизма, происходит запуск адаптивных систем, нивелирующих нарушения, возникающие в результате воздействия ионизирующего излучения на организм.А. И. Дворецким (1986) предложена гипотеза вероятной последовательности процессов нарушения энергетических и регуляторно-транспортных функций мембран в условиях воздействия ионизирующей радиации.
Инициирующим процессом при действии лучевого фактора является активация пероксидного окисления липидов в клетке, происходящая на фоне резкого усиления эндогенного фосфолипазного гидролиза. В результате накопления продуктов ПОЛ происходит изменение баланса между эндогенными радиопротекторами и радиосенсибилизаторами в пользу последних.
Окисление мембранных липидов и повышенный эндогенный фосфолипазный гидролиз приводят к нарушению проницаемости биомембран для катионов. Это вызывает уменьшение уровня калия, увеличение содержания натрия и возрастание концентрации кальция в цитозоле, поэтому происходит насос-зависимое увеличение объема клетки, нарушение структурно-функционального состояния Na+, К?-АТФазы за счет конформационных изменений молекул фермента на фоне липид-белковых перестроек в мембране. В ответ на изменение ионного гомеостаза возможно компенсаторное увеличение количества молекул Na+, К+-АТФазы за счет “резервных”. Однако вследствие деэнергизации клетки, индуцированной нарушением окислительного фосфорилирования и дефицита АТР, функционирование Na+, К+-АТФазы оказывается затрудненным. Изменяется и активность Са2+-АТФазы. Все это, наряду с нарушением пассивной проницаемости, вызывает резкое увеличение уровня внутриклеточного кальция. В результате выявляются необратимые последствия, связанные с модификацией мембранного скелета и изменением формы клетки. Существенным моментом в патофизиологии клеточной гибели является вторичная активация фосфолипаз кальцием. Это приводит к фосфолипазному истощению мембран клеток и внутриклеточных органелл и возрастанию их проницаемости. Нарушения в функционировании всех клеточных систем способны индуцировать необратимые изменения в процессе жизнедеятельности клеток.
Необходимо отметить, что компоненты биологических мембран играют ключевую роль в развитии свободнорадикальных реакций в клетках и тканях, индуцируемых воздействием ионизирующего излучения, и патогенезе лучевого поражения живых организмов. Процессы нарушения липидного состава, текучести липидных компонентов, подвижности липидных и белковых молекул, липид-белковых взаимодействий, изменения активности мембраносвязанных ферментов, собственных систем антиради- кальной защиты представляют собой комплексный и взаимосвязанный механизм, индуцирующий пострадиационные структурно-функциональные перестройки биомембран и обеспечивающий переход клеток и организма на новый метаболический уровень функционирования.
4.2.