Гормоны насекомых и феромоны
У насекомых контроль большинства физиологических процессов осуществляется специфическими гормонами. Изучение этих гормонов с точки зрения избирательной токсичности имеет две цели: во-первых, изучение природы гормонов и разработка экономичных методов их синтеза для применения в количествах, обеспечивающих нарушение метаболизма у насекомых; во-вторых, синтез аналогов, обладающих антагонистическим действием (разд.
9.1), для использования их в качестве инсектицидов.Специфические гормоны насекомых следующие: а-экдизон, гормон линьки, образующийся в железах, расположенных в переднегруди насекомых; гормон мозга, стимулирующий деятельность желез переднегруди; ювенильный гормон, вырабатываемый в corpora allata и стимулирующий метаморфоз; гормон, выделяемый corpus cardiacum, увеличивающий амплитуду сокращений сердечной мышцы и перистальтику кишечника, и ади- покинетический гормон, регулирующий метаболизм липидов •—• главного источника энергии для полета [Stone et al., 1976].
Экдизон (4.77) обладает некоторыми особенностями структуры, не встречающимися у стероидных гормонов млекопитающих: кольца А и В сконденсированы в цис-положении, у атома С-2 находится гидрофильный заместитель, кроме того, в а-части цепи (Сз—Ci5) также содержатся гидрофильные заместители, а у Сп — очень длинная боковая цепь (подробнее о стероидах
см. разд. 12.2). Существенной для наличия активности является кето-группа в положении 6. Крустэкдизон, гормон линьки ракообразных, встречающийся также в некоторых насекомых, представляет собой 20-гидроксиэкдизон. Соединения группы экдизо- нов можно получать в достаточных количествах из растительных продуктов. Несмотря на то что при инъекционном введении многие аналоги экдизона обладают инсектицидной активностью, они практически не находят применения из-за того, что не проникают через кутикулу насекомых.
Сами насекомые не могут синтезировать эти гормоны, как и любые другие стероиды, de novo, и вынуждены получать с пищей промежуточные для их синтеза продукты [Lasser, 1966]. Эта особенность насекомых — мишень для избирательного воздействия.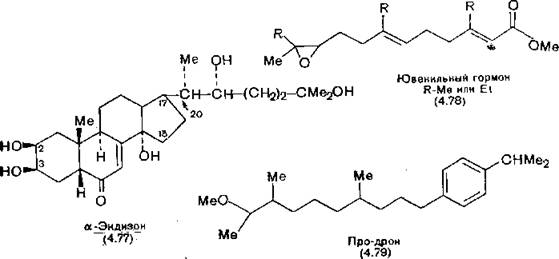
Ювенильный гормон (4.78)—простое по структуре алифатическое соединение — метиловый эфир транс, транс-3,11-диметил-10-эпокси-7-этилтридека-2,6-диеновой кислоты [Roller et al., 1967]. Активность его в основном определяется конфигурацией С-1 конца молекулы.
При создании миметиков (агонистов) ювенильного гормона было установлено, что нестабильность подобных соединений связана с наличием эпоксидного цикла. Поэтому в дальнейшем он был исключен из молекулы. Типичные миметики на конце цепи рядом с двойной связью чаще всего содержат эфирную группу, находящуюся в транс-положении по отношению к алкильной цепи. Таково, например, строение метопрена — изопропилового эфира (2Е, 4Е)-11-метокси-3,7,11-триметилдодека-2,4- диеновой кислоты. Метопрен используют для борьбы с москитами и навозными мухами, его применяют в виде полиамидных гранул, из которых он медленно диффундирует. Основное действие агонистов ювенильного гормона заключается в том, что они задерживают развитие насекомых на стадии личинки, наносящей наибольший вред. Поэтому эти миметики применяют главным образом для борьбы лишь с теми видами насекомых,
у которых наибольший вред причиняют именно взрослые особи, например москитами и мухами. Структурный аналог ювенильного гормона «Про-дрон» (4.79) отличается от гормона наличием в молекуле бензольного кольца и отсутствием двойных связей и обладает совершенно другим типом действия [Schwarz, Miller, 1979]. Хотя это соединение вначале испытывали как средство борьбы с комнатными мухами, оно оказалось эффективным против муравьев Рихтера. Попав с пищей в передний желудок муравьев, оно сохраняется там в течение нескольких месяцев и в результате число рождений трутней (самцов) превышает число рождений рабочих особей, вследствие чего муравьи вымирают от голода.
Никаких побочных эффектов для людей, птиц и рыб у этого соединения не обнаружено.Типичными представителями антагонистов ювенильного гормона являются прецоцены, например (4.80) —>хромон, содержащийся в садовых растениях. Даже в маленьких концентрациях они вызывают преждевременную линьку, приводящую к стерильности взрослых особей [Bowers et al., 1976]. Место их действия— железа, синтезирующая ювенильный гормон, который активирует молекулы прецоценов, по-видимому, эпоксидированием. Эти эпоксиды, в свою очередь, разрушают сам ювенильный гормон [Brooks, Pratt, Jennings, 1979].
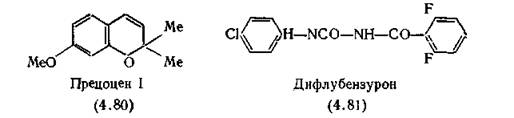
Один из наиболее широко применяемых антагонистов гормонов насекомых дифлубензурон (4.81) [3-(2,6-дифторбензоил)-1- (4-хлорфенил)мочевина], ингибирует образование фермента, превращающего УДФ-ацетилхитозамин в хитии, прекращая тем самым линьку мух и москитов. Кроме того, он блокирует созревание яиц и поэтому такие насекомые, как мухи цеце, откладывающие живые личинки, не дают жизнеспособного потомства. Для человека дифлубензурон безвреден [Jordan, Trevern, 1978].
Опубликованы обзоры по физиологии гормонов насекомых [Novak, 1975] и по биохимии насекомых k[Candy, Kilby, 1975].
Феромоны насекомых — средства информации между особями — выделены из многих видов. Эти высокоспецифичные соединения эффективны даже в следовых количествах. Чаще всего это алифатические соединения, как, например, додека- триен-1-ол — вещество, которым термиты отмечают свой путь. Однако городские муравьи секретируют метиловый эфир 4-ме- тилпиррол-2-карбоновой кислоты, а феромоны некоторых муравьев и жуков являются производными пиразина [Barlin, 1982].
Феромоны другого типа — половые аттрактанты — секретируются только самками. Самка тутового шелкопоряда выделяет 10-транс-12-цис-гексадека-10,12-диен-1-ол, самки непарного шелкопряда— цис-2-метил-7,8-эпоксиоктадекан, а самки бабочки хлопковой совки— 10-пропилтридека-5,9-диенол.
«Королевская субстанция», которая выделяется железой, расположенной в нижней челюсти пчелы-матки, и останавливает развитие личинок пчел на стадии рабочих особей, представляет собой транс-9-оксо-дека-2-еновую кислоту: СН3СО(СН2)5СН=СНСООН.
Феромоны насекомых применяют как в качестве приманок к отравленным ловушкам, так и с целью нарушения полового поведения насекомых. Однако их применение ограничено прежде всего высокой специфичностью к определенному виду насекомых, а также их высокой стоимостью. Подробнее о феромонах см. Rothschild (1981) и Kydonieus, Beroza (1982).
Процесс развития самцов пустынной саранчи ускоряется, если насекомые собираются в стаю. Можно предполагать, что этому способствуют летучие стимулянты.
4.7.3.
Еще по теме Гормоны насекомых и феромоны:
- Феромоны и гормоны растений
- Гормоны и феромоны
- Пол и гормоны
- Гормоны позвоночных
- Глава шестая: «Сигнал весны на языке гормонов»
- 4.1.3.1. Класс Insecta (насекомые)
- Ядовитые насекомые
- Класс Насекомые
- Вещества, действующие на нервую систему насекомых
- В виде насекомых (2)
- Глава XXXII Ужаление пчелами и насекомыми
- Надотряд Holometabola (насекомые с полным метаморфозом)
- Надотряд Hemimetabola(насекомые с неполным метаморфозом)