2.1.2. Рецепторы на ферментах
Вернемся к блокированию АХЭ физостигмином — первому примеру взаимодействия лекарственного вещества с рецептором на ферменте (разд. 2.1.0). Изучение аналогов физостигмина пока-
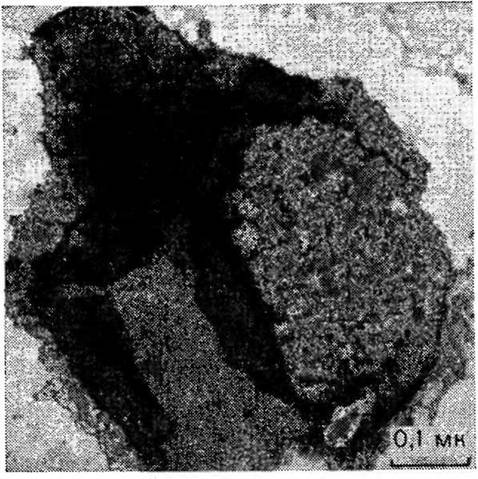
Рис.
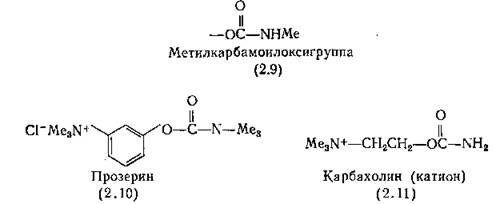
2.1. Электронная микрофотография плазматической мембраны электрического органа рыбы (Torpedo) с расположенными на ней рецепторами АХ. Центр каждой розетки — Na/K+ канал.зало, что для связывания с АХЭ необходимо наличие в молекуле двух фрагментов: основной группы и метилкарбамоилокси- группы (2.9) [Stedman, 1926; Stedman, Stedman, 1931]. Прозе- рин (2.10), простейший из аналогов физостигмина, оказался самым активным препаратом при лечении миостении гравис, болезни, проявляющейся в слабости различных групп мышц и по симптомам напоминающей отравление тубокурарином. Как и физостигмин, прозерин содержит метилкарбамоильную группу,, а его основная группа такая же, как у АХ. Наличие этих двух групп позволяет прозерину легко вытеснять АХ из активного центра АХЭ, но в отличие от физостигмина он может проявлять и прямое агонистическое действие на мышечный холинорецептор.
Основанное на этих данных предположение о сходстве кар- бамоилоксигруппы («уретановой») с ацетильной группой аце-^ тилхолина привело к синтезу карбахолина (карбамоилхолина) (2.11) , обладающего аналогичной АХ активностью, но действующего более продолжительно [Molitor, 1936]. Хотя частично его действие истинно агонистическое (он взаимодействует с постсинаптическими рецепторами), тем не менее преимущественно оно псевдоагонистическое: карбахолин способствует высвобождению АХ из пресинаптического нервного окончания, что Резко отличает его от физостигмина. Действие карбахолина не блокируется, а сам он не гидролизуется АХЭ. Препарат приме-
няют для восстановления тонуса мочевого пузыря и кишечника после операций.
Из фармакодинамических лекарственных средств к ингибиторам ферментов относятся и сердечные гликозиды, ингибирующие Na—К АТФазу (см. разд. 14.1) [Bonting, 1970].
До 1940 г. фармакодииамика и химиотерапия развивались независимо друг от друга, пока D. Woods не показал, что про- тивобактериальные свойства стрептоцида (2.13) связаны со структурным и электронным сходством строения его молекулы с молекулой пара-аминобензойной кислоты (ПАБ) (2.12) [Woods, 1940]. Позднее это количественно подтвердили Bell, Roblin (1942). При одновременном изменении стерических и электронных свойств молекулы стрептоцида, приводящем к увеличению его сходства с ПАБ, противобактериаль- ная активность вещества усиливается, если же при изменении свойств сходство уменьшается, то активность ослабляется. Механизм биологического действия сульфаниламидов установил Gene Brown, выделив фермент дигидрофолатсинтетазу, синтезирующую важный кофермент — дигидрофолиевую кислоту (2.14), составной частью которой является ПАБ. В молекуле дигидро- фолатсинтетазы есть место специфического связывания (рецептор) ПАБ. При высоких концентрациях сульфаниламид, согласно закону действующих масс, вытесняет природный субстрат с рецептора на ферменте, что приводит к ингибированию действия фермента и нарушению синтеза дигидрофолиевой кислоты [Brown, 1962].
Противобактериальные сульфаниламиды, действие которых после их открытия в 1930 г. считали основным доказательством гипотезы «одна группа — одно действие», в конце концов нанесли ей смертельный удар. Стало очевидным, что наличие в молекуле сульфамидной группы еще не определяет ее противобакте- риальную активность, и для взаимодействия молекулы препарата с рецептором необходимо соблюдение и других структурных требований (например, наличие заместителя в пара-положении, см. разд. 9.3.1). При выполнении этих структурных требований сульфаниламидную группу можно заменить на любую другую со сходной полярностью.
Например, более выраженными про- тивобактерикльными свойствами, чем стрептоцид, обладает 4,4/-диаминооензил (2.15), также ингибирующий связывание ПАБ с ферментом [Kahn, Weygand, МбНег, 1943]. С другой стороны, ароматическая сульфаниламидная группа входит в молекулы многих противодиабетических лекарственных веществ и диуретиков, такиД как, например, бутамид (12.61) и хлортиазид (14.1) соответственно. Однако размеры этих молекул и распределение в них зарядов делает невозможным их взаимодействие с дигидрофолатсинтетазой, поэтому они не обладают проти- вобактериальным действием. В то же время блокаторы этого фермента, обладающие противобактериальным действием, могут вообще не иметь в своей молекуле атома серы.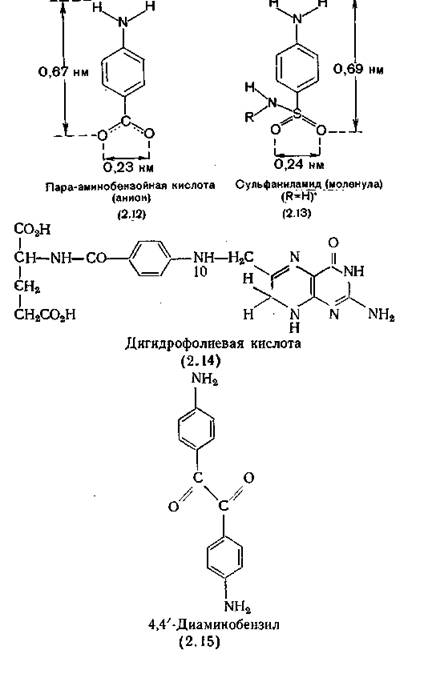
И для других лекарственных веществ существуют рецепторы ■на ферментах, например для антибиотика рифампицина (4.37), ингибирующего действие РНК-полимеразы (раз^/4.0). Аллопу- ринол (9.51), специально созданный для того, Чтобы блокировать два взаимозависимых фермента — гипокрантиноксидазу и ксантиноксидазу [Elion et al., 1966], ингибирует образование мочевой кислоты и поэтому успешно используется для лечения хронической подагры (разд. 9.4.4). Противомалярийное действие хлоридина и противобактериалыюе действие триметоприма связаны с их блокирующим действием на фермент дигидрофолатре- дуктазу (разд. 9.3.3). Противобактериальная активность пенициллина проявляется в ингибировании полимеразы, участвующей в образовании новой клеточной стенки бактерий (разд. 13.1). В главе 9 приведены примеры лекарственных веществ, рецепторы которых расположены на ферментах.
Идея существования рецепторов для инсектицидов на ферментах нашла подтверждение вскоре после второй мировой войны, когда стали применять фосфорорганические вещества, ингибирующие АХЭ насекомых этерификацией гидроксильной группы остатка серина, входящего в состав этого фермента (разд. 13.3).
Действие ферментов на молекулярном уровне обсуждается в разд. 9.0.
2.2.