Активация лимфоцитов
Воспаление может рассматриваться как элемент общего иммунного ответа организма на чужеродное воздействие, при этом центральным звеном противодействия антигенной агрессии выступают Т- и В-лимфоциты.
Клональная теория иммунного ответа предполагает, что в результате контакта с антигеном, расположенным на поверхности антиген-представляющей клетки, индуцируется пролиферация определенного клона Т-хелперных лимфоцитов, которые в свою очередь вызывают дифференцировку В-лим- фоцитов в продуцирующие антитела плазматические клетки. Такая функциональная кооперация клеток в иммуногенезе включает разные костимулирующие сигнальные пути, начиная от CD-рецепторных контактов, обеспечивающих физическое сближение клеток, и заканчивая синтезом широкого спектра цитокинов, регулирующих рост и дифференцировку клеток. Наработка АКМ мембрансвязанными NADPH-оксидазами в фагоцитирующих клетках важна для микробицидного действия, однако АКМ также могут участвовать в развитии гуморального иммунного ответа посредством индукции пролиферации и дифференцировки лимфоцитов [596, 1403]. Сегодня достаточно убедительно показано, что митогенная стимуляция лимфоцитов сопровождается увеличением продукции супероксидного анион-радикала, который необходим для индукции пролиферации клеток, и его удаление ингибирует реакцию лимфоцитов на пролиферативные стимулы [687, 1403].Среди продуктов неполного восстановления молекулярного кислорода (О2, НО’, Н2О2, ОН’) по ряду физико-химических свойств именно молекула H2O2 наиболее удачно подходит на роль вторичного мессенджера для активации лимфоцитов [1403]. Во-первых, H2O2 более стабильна относительно O2 и ОН-радикалов, время ее жизни определяется наличием восстанавливающих ферментов (каталазы, глутатионпероксидазы и др.) и в биологических средах составляет около 1 мс, что достаточно для межклеточной диффузии.
Молекула не имеет заряда и хорошо проникает через мембраны. В реакционном плане Н2О2 существенно менее агрессивна по сравнению с ОН-радикалами, поэтому она избирательно и, как правило, обратимо окисляет цистеиновые аминокислоты в составе белков. Цистеин хорошо взаимодействует с Н2О2 только когда присутствует в форме цистеинтиолат-аниона (—СН2—8_), для большинства цистеиновых остатков в белках рКа около 8,5, и при физиологических рН сера связана с водородом. Однако если цистеин соседствует с положительно заряженными аминокислотными остатками, то рКа может снижаться до 5,0, в этом случае цистеин депротеинируется и легко окисляется перекисью водорода в сульфеновую кислоту, которая в последующем подвергается нуклеофильной атаке с образованием либо дисульфидных, либо сульфениламидных связей [1575].Несмотря на то, что посредством ингибирования NAD^^^^na^ и исследования животных, нокаутированных по ее различным субъединицам, достаточно убедительно показано участие активных форм кислорода в индукции пролиферации лимфоцитов, механизм данного действия остается неясным. Предполагается, что действие Н2О2 на лимфоциты может реализоваться через редокс-чувствительные факторы транскрипции (р53, NF-κΒ, АР-1, HIF и др.), пероксиредоксины или регуляцию фосфорилирования белков [160, 665, 1342, 1575]. В последнем случае особого внимания заслуживает регуляция протеинтирозинфосфатаз, которые участвуют в регуляции многих ферментов посредством их дефосфорилирования [1637]. Протеинтирозинфосфатазы в активном центре содержат редокс-чувствительный цистеин, окисление которого перекисью водорода переводит фермент в неактивное состояние (рис. 4). Действие АКМ на лимфоциты может также реализоваться через активацию специфических протеинтирозинкиназ, действующих на факторы транскрипции [1689].
Индуцирующая пролиферацию и дифференцировку лимфоцитов перекись водорода может быть экзогенного (антиген-представляющие клетки) или эндогенного происхождения.
Действительно, в Т-лимфоцитах показано наличие мембранной Са2+-зависимой Nox5, продукция О2 которой индуцирует пролиферацию, при этом индуцированная конканавалином А пролиферация ингибируется дифенилениодони-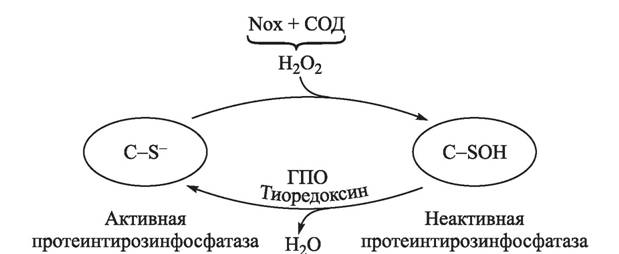
Рис. 4. Регуляция активности протеинтирозинфосфатаз H2O2 [1403]
ем (эффективный ингибитор NAD(P)H-оксидаз) [174, 647]. Активация Т-лимфоцитов антиген-представляющими клетками ингибировалась полифенолами (кверцетин и рутин), а также N-ацетилцистеином [671]. Вместе с тем обработка Т-лимфоцитов человека специфическим ингибитором γ-глутаминцистеинсинтетазы L-бутионин-(S,R)- сульфоксимином приводила к снижению содержания внутриклеточного глутатиона и ингибированию пролиферативного ответа на антигены [1587]. Индуцированная бактериальным липополисахаридом или мембранным иммуноглобулином пролиферация и дифференцировка мышиных В-лимфоцитов ингибировалась антиоксидантами бутил- гидрокситолуолом, орто-фенантролином, тиомочевиной и α-токоферолом [596].
У животных, нокаутированных по фактору транскрипции Nrf2, выявляются множественные аутоиммунные воспалительные процессы, которые характеризуются высокой пролиферативной активностью СD4+ Т-лимфоцитов [1064]. Известно, что антиоксидант- респонсивный элемент и фактор транскрипции Nrf2 запускают синтез ферментов антиоксидантной защиты и метаболизма ксенобиотиков при различных стрессовых воздействиях, в том числе и окислительном стрессе [131]. По-видимому, активация антиоксидант-рес- понсивного элемента экзогенными антиоксидантами может лежать в основе изменения поляризации макрофагов и ингибирования воспалительного процесса [79, 80].
Пролиферация (от лат. proles — «потомство» и fero — «нести») и апоптоз — важные элементы воспаления, определяющие завершение процесса и восстановление гомеостаза. Если на начальных стадиях воспаления преобладают деструктивные процессы, то на более поздних стадиях усиливается клеточная пролиферация, индуцируется синтез коллагена, что необходимо для восстановления структуры и функции органа.
Процесс пролиферации находится под сложным гуморальным контролем. Макрофаги и лимфоциты синтезируют целое семейство цито- и лимфокинов, многие из которых, и прежде всего фактор роста фибробластов, оказывают влияние на деление клеток. В настоящее время накопилось много данных, показывающих участие АКМ и радикальных окислительных реакций в клеточной пролиферации, опухолевой трансформации и гибели клеток [429]. Развитие окислительного стресса при воспалении одновременно является сигналом для клеточной пролиферации и апоптоза определенных клеточных клонов.Первая публикация, демонстрирующая стимуляцию роста клеток экзогенными прооксидантами, появилась в 1947 г. [370]. Последующие исследования показали, что антиоксиданты, в том числе витамин Е, обладают выраженным антипролиферативным действием [437]. Считается, что окислительный стресс средней интенсивности, вызванный супероксидным анион-радикалом и перекисью водорода, стимулирует пролиферацию, в то время как высокие концентрации АКМ инициируют гибель клеток посредством апоптоза (физиологически достигаемые концентрации АКМ) или некроза (при концентрациях выше апоптогенных значений). Чувствительность разных типов клеток к экзогенным АКМ зависит от антиоксидантной защиты и активности систем репарации и существенно различается. Развитие окислительного стресса в легких может индуцировать апоптоз пневмоцитов II типа, но усиливать пролиферацию фибробластов, что является одной из причин фиброза [1198].
Зрелые гранулоциты, циркулирующие в крови, уже имеют начальные морфологические признаки апоптоза (фрагментация ядра, конденсация хроматина), процесс их гибели развивается необратимо в течение 2—3 суток. Гранулоциты первыми поступают в очаг воспаления, где активируются и после гибели (преимущественно посредством апоптоза) удаляются макрофагами. Усиление продукции АКМ и развитие окислительного стресса предшествует апоптозу гранулоцитов. В условиях гипоксии, а также при генетических нарушениях способности генерации АКМ NADPH-оксидазой (больные хроническим гранулематозом) запрограммированная гибель нейтрофилов снижена [857].
При этом как повышенная, так и сниженная способность гранулоцитов к генерации АКМ и апоптозу может служить причиной хронизации воспалительного процесса, в первом случае экссудативно-деструктивного типа, во втором — мононуклеарного.Воспалительный процесс в тканях сопровождается значительной локальной продукцией АКМ, и прежде всего H2O2 как наиболее стабильной формы АКМ. Среди разных типов лимфоцитов наименее чувствительны к цитотоксическому действию перекиси водорода Т-хелперы (CD4+ Т-лимфоциты), что служит причиной увеличения соотношения Т-хелперы/Т-супрессоры в очаге воспаления при действии активированных нейтрофилов [1849]. У ВИЧ-инфицированных людей отмечается падение содержания Т-хелпер- ных лимфоцитов в результате их апоптоза. Исследования инфицированных вирусом клеток показали, что в них на 50 % уменьшено содержание каталазы [1453], а также снижено содержание другого ингибирующего H2O2 белка — тиоредоксина [1113]. При этом клетки становятся чувствительными к действию H2O2, их саморазрушение начинается при концентрации перекиси в культуральной среде больше 10 мкМ. Таким образом, ослабление антиоксидантной защиты против H2O2 в Т-хелперных лимфоцитах при ВИЧ-инфекции приводит к уменьшению размеров их популяции и развитию иммунодефицитного состояния. Аналогичная, но менее выраженная, повышенная способность Т-хелперов к апоптозу выявлена у людей, инфицированных вирусом герпеса [1808].
Проблема клеточной пролиферации сложна, и ее нельзя решить только поиском новых факторов роста; по-видимому, существуют более общие древние механизмы регуляции размножения клеток. В процессе эволюции одним из «исторических моментов» зарождения таких механизмов следует считать появление митохондрий в результате симбиоза эукариотической и прокариотической клеток. Если в каждой из клеток была заложена программа на рост и деление, то жизнеспособность симбионта была возможна только в результате согласования этих программ.
Можно предположить гипотетическую схему такого согласования через наработку продуктов неполного восстановления молекулярного кислорода (АКМ), которая затем трансформировалась в один из механизмов регуляции клеточной пролиферации и апоптоза, в частности, в очаге воспаления. Существует мнение, что в физиологических условиях в результате апоптоза из популяции удаляются клетки с аномально высоким уровнем продукции АКМ [194], что снижает риск опухолевой трансформации клеток. Более подробно участие АКМ в клеточных процессах рассмотрено в разделах, посвященных внутриклеточному окислительному стрессу и апоптозу.Таким образом, можно видеть, что АКМ активно участвуют в развитии воспалительной реакции, начиная с фазы альтерации и кончая процессами репарации. При этом, как отмечалось выше, регуляторное, микробицидное и деструктивное действие АКМ не ограничено рамками иммунной системы или микроциркуляторного русла. Все клетки организма имеют механизмы синтеза АКМ, а следовательно — цитотоксического и регуляторного действия. При этом свободнорадикальная концепция позволяет объединить клеточную, сосудистую и нейроиммунную теории воспалительного процесса и органически рассмотреть проблему термогенеза (calor), которая не может быть решена без рассмотрения радикальных окислительных реакций, являющихся ключевым звеном процесса теплопродукции в клетках [537].