§ 3. Способна ли миграция стабилизировать экосистему?
В предыдущем параграфе мы имели пример того, как неодинаковость параметров среды в разных точках пространства ведет к потере устойчивости равновесных состояний. Возникает вопрос, а каково влияние пространственно распределенной структуры на устойчивость, если значения параметров среды одинаковы? Мы попытаемся дать ответ на этот вопрос, рассмотрев динамику нескольких одинаковых сообществ, связанных между собой миграционными потоками.
Допустим, что в пространстве существует некоторое число п одинаковых, относительно изолированных местообитаний, в каждом из которых живет сообщество с одним и тем же набором р видов, взаимодействующих между собой одинаковым образом во всех местообитаниях. Допустим также, что особи видов, принадлежащих отдельным сообществам, могут, в соответствии с их радиусами индивидуальной активности, совершать переходы из одних местообитаний в другие, т. е. что происходит миграция видов между сообществами. Естественно считать, что интенсивность миграции не безразлична к численностям мигрирующего вида в тех местах, откуда и куда происходит миграция.
Ясно тогда, что совокупность п сообществ образует уже целую систему — будем называть ее объединенной, — состоящую из п подсистем. Внутри подсистемы виды связаны биологическими взаимодействиями, а между подсистемами идут потоки мигрантов. Можно ли ожидать устойчивости в объединенной системе, если устойчивостью обладают изолированные подсистемы? Способны ли миграционные связи стабилизировать неустойчивое поведение изолированных подсистем? На эти вопросы экологи склонны давать утвердительный ответ, а что дает анализ математических моделей, станет ясно из последующего изложения.
Пусть динамика изолированного сообщества моделируется некоторой системой уравнений
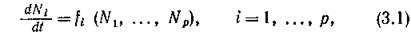
где на правые части наложены лишь качественные ограничения существования равновесия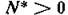 и допустимости
и допустимости
линеаризации в точке Объединяя п таких подсистем, верхним индексом при переменной
Объединяя п таких подсистем, верхним индексом при переменной будем указывать номер соответствующей подсистемы.
будем указывать номер соответствующей подсистемы.
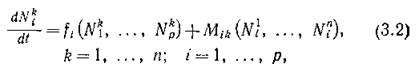
где функции fi задают структуру биологических взаимодействий внутри /г-й подсистемы (достаточно произвольную, но одинаковую во всех подсистемах), а функции M(k описывают влияние миграционных потоков из остальных подсистем в/г-ю. Простейший вид этих функций — линейный — соответствует миграционным потокам с интенсивностями, постоянными для і-го вида и для фиксированного маршрута миграции (т. е. направления Разумеется, линейная зависи
Разумеется, линейная зависи
мость — далеко не единственный тип зависимости миграции от численностей мигрирующих видов, однако поскольку все дальнейшие результаты получены нами методом линеаризации, вполне можно ограничиться линейным приближением.
Если допустить, что схема миграционного процесса (маршруты мигрантов между подсистемами) определяется только геометрией взаимного расположения местообитаний в пространстве (или, иными словами, географическими особенностями), то ясно, что эта схема должна быть одинакова для всех мигрирующих видов. В этом случае она предста-
Замкнутость системы по миграции означает, что
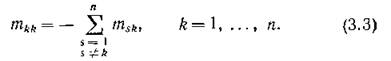
Обозначая через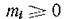 ту долю интенсивности мигра
ту долю интенсивности мигра
ции, которая определяется только биологическими особенностями і-го вида (і = 1, ... , р) и не зависит от маршрута миграции, получаем систему уравнений объединенного сообщества в виде 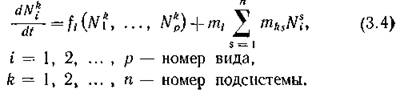
Равенство ті = 0 означает, что і-й вид (во всех подсистемах) не участвует в миграции.
Если интенсивность миграции целиком определяется лишь биологическими особенностями видов и не зависит от направления (среда изотропна по миграции), то все ненулевые элементы матрицы Мп, не стоящие на главной диагонали, одинаковы и лишь указывают на то, что между соответствующей парой местообитаний происходит миграция (с одинаковой в обоих направлениях интенсивностью). В таком случае все эти элементы можно положить равными 1; матрица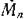 симметрична, и в силу (3,3) все ее строчные муммы равны нулю:
симметрична, и в силу (3,3) все ее строчные муммы равны нулю: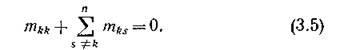
Легко видеть, что тогда уравнения стационарного состояния системы (3.4)
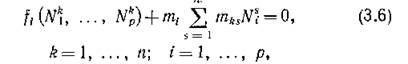
допускают решение в виде лр-вектора
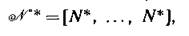
в котором сохраняются все прежние равновесные значения численностей изолированных подсистем [20]). Линеаризация (3.4) в данной точке дает матрицу блочно-диагональной структуры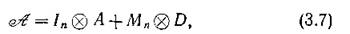
где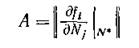
— матрица линеаризованной изолированной подсистемы p-го порядка,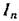 — тождественная матрица «-го порядка,
— тождественная матрица «-го порядка,
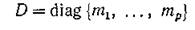
— диагональная матрица интенсивностей миграций, зависящих лишь от биологических особенностей каждого вида,

деленным образом сужает (либо делает пустой) область устойчивости в пространстве параметров модели.
Если спектр А — вещественный, а среди дополнительных значений, возникающих в спектре есть комплексные, то монотонный характер траекторий в окрестности
есть комплексные, то монотонный характер траекторий в окрестности
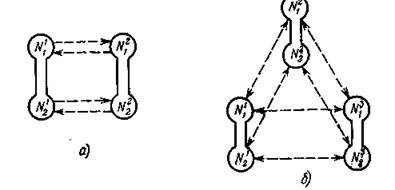
Рис. 44. Примеры миграционных схем. Штриховые стрелки соответствуют потокам мигрантов; взаимодействие внутри местообитаний не конкретизируется.
равновесия изолированной подсистемы меняется на колебательный. Противоположной смены произойти, очевидно, не может, поскольку сохраняется спектр А.
Пример 1. Пусть п = р = 2 (см. рис. 44, а). Тогда объединенная система имеет четвертый порядок:
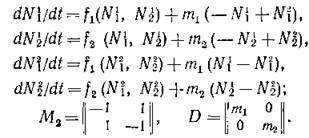
Обозначив элементы матрицы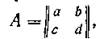
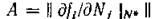 через
через
получаем матрицу объединенной системы 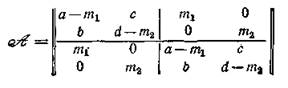
с характеристическим уравнением
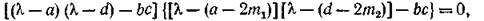
имеющим, наряду с корнями изолированной подсистемы
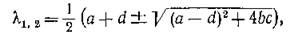
пару новых корней
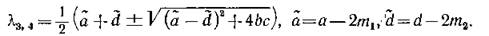
Критерий устойчивости по линейному приближению дает условия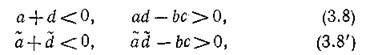
при выполнении которых все Re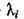 отрицательны.
отрицательны.
Ясно, что если выполняются неравенства (3.8) — усло
вия устойчивости изолированной подсистемы, — то при достаточно малых тл и т2 выполняются и (3.8'), т.
е. устойчива и объединенная система. Если траектории изолированной подсистемы в окрестности N* монотонно устойчивы, что имеет место, когда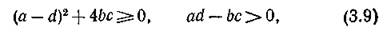
то монотонность устойчивых траекторий объединенной системы сохранится, если
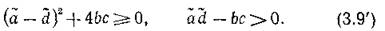
Когда соблюдено условие (3.9), при достаточно малых т1 и т2, очевидно, выполняется и (3.9'). Если же т± и ms таковы, что нарушается первое из условий(3.9'), т. е. возникают комплексно-сопряженные корни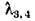 и выполнено условие
и выполнено условие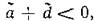 то монотонный характер устой
то монотонный характер устой
чивых траекторий изолированной подсистемы меняется на колебательно-устойчивое поведение траекторий объединенной системы. Противоположной смены произойти не может из-за наличия в спектре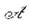 пары корней Х1>2 изолированной подсистемы, т. е. если в подсистеме наблюдались колебания при стремлении к равновесию
пары корней Х1>2 изолированной подсистемы, т. е. если в подсистеме наблюдались колебания при стремлении к равновесию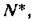 то колебания сохранятся и после объединения в систему.
то колебания сохранятся и после объединения в систему.
Аналогичным образом могут быть рассмотрены и условия, при которых монотонная неустойчивость приобретает после объединения колебательный характер.
Если же при выполнении (3.8) второе из неравенств (3.8') меняется на противоположное (первое, очевидно, нарушиться не может), равновесие объединенной системы оказывается неустойчивым, т. е. миграция дестабилизирует систему.
Пример 2. Другой пример миграционной структуры представлен на рис. 44, б, где п = 3. Здесь с одинаковыми интенсивностями миграция происходит между любой парой из п местообитаний, что можно объяснить равноудаленностью их друг от друга на плоскости.
В трехмерном пространстве аналогичные геометрические соображения приводят к четырем местообитаниям, расположенным в вершинах правильного тетраэдра. При п > 4 подобные геометрические представления в трехмерном пространстве уже невозможны.Матрица миграционной структуры (в изотропном случае) имеет здесь вид
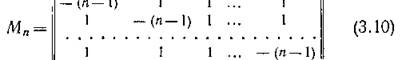
и удовлетворяет условиям леммы 1.
Вообще, можно представить и более сложные схемы
объединения подсистем, чем изображенные на рис. 44, а, б. Лемма 1 позволяет утверждать, что прежние собственные числа сохраняются в спектре объединенной системы при любой миграционной структуре Мп, имеющей нулевые строчные суммы. Так как последнее условие является и критерием сохранения прежних равновесных значений в стационарном решении объединенной системы (3.6), отсюда следует такой вывод: если в результате объединения по какой-либо схеме миграции сохраняются равновесные значения изолированных подсистем, то и в спектре объединенной системы сохраняются прежние собственные числа и справедливы сформулированные выше утверждения о характере траекторий в окрестности равновесия.
В частном случае, когда все «биологические» интенсивности миграции одинаковы, т. е. 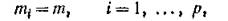
техника кронекеровских произведений матриц позволяет получить явный вид всех собственных чисел объединенной системы. Действительно, в этом случае

Пример 3. Для схемы миграции, возникающей при «кольцевом» расположении местообитаний, аналогичном кольцевому расположению экологических ниш конкурирующих видов (см. рис. 39 гл. VI), матрица миграционной структуры в изотропной среде представляет собой циркулянт
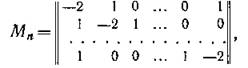
собственные числа которого равны
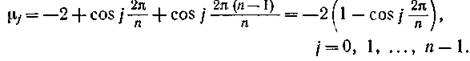
В частном случае одинаковости всех mi = т собственные числа равны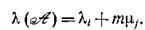
Наличие [х0 = 0 подтверждает вывод о сохранении всех прежних собственных чисел и поскольку все остальные р,- вещественны и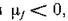 можно сказать, что объеди
можно сказать, что объеди
нение не ухудшает свойств устойчивости по сравнению с изолированным случаем и не изменяет качественной картины поведения траекторий в окрестности равновесия.
Подводя итог рассмотрению миграционных схем в изотропной среде, мы убеждаемся (лемма 1), что при любой структуре биологических взаимодействий в изолированной подсистеме объединение посредством миграции в изотропной среде не способно улучшить свойств системы с точки зрения устойчивости, а может лишь изменить монотонное поведение траекторий в окрестности равновесия (устойчивого либо неустойчивого) на колебательное, но не наоборот. В некоторых случаях такое объединение может даже дестабилизировать устойчивые подсистемы (см. пример 1).
Основанием этих математических результатов служит, как было показано, вырожденность матрицы миграционной структуры Мп для изотропной среды. В то же время трудно представить, чтобы любая реальная среда, где практически всегда имеются какие-то преимущественные направления миграции, обеспечивала бы строгую изотропность на всех маршрутах. По-видимому, стабилизирующие эффекты миграции, наблюдаемые в экспериментах, могут быть объяснены неодинаковостью интенсивностей миграции в различных направлениях, а в рамках математических моделей подобные эффекты следует искать лишь в таких схемах, где интенсивности существенно зависят от направления, т. е. среда анизотропна по миграции. Рассмотрению такого рода схем и посвящены два следующих параграфа.
Еще по теме § 3. Способна ли миграция стабилизировать экосистему?:
- Деятельностный подход к анализу и объяснению способностей. Способности и задатки
- § 4. Миграция в анизотропной среде
- 7. Незаконная миграция населения
- § 1. Институт миграции: понятие и сущность
- § 1. Понятие международной трудовой миграции
- Миграция населения
- 3.4. Миграция рабочей силы и её регулирование
- 30 Миграционная политика. Регулирование внешней трудовой миграции.
- § 5. Метод малого параметра в исследовании эффектов миграции
- Тема 3. Миграция населения России
- Международная миграция капитала: сущность, причины и формы
- § 2. Миграция в Европейский Союз в свете принципов и норм международного права
- Регулирование международной миграции рабочей силы
- 60-61. Интернациональные экономические отношения [1]- Миграция капитала и рабочей силы.
- Сколько было волн миграции и откуда они?
- 29 Международная миграция рабочей силы: понятие, причины, элементы, современные тенденции развития.