§ 4. Миграция в анизотропной среде
Перейдем теперь к рассмотрению случая, когда интенсивность миграции зависит не только от биологических особенностей мигрирующих видов, но и от маршрута, направления миграции, иными словами, будем считать, что среда анизотропна по миграции.
Влияние анизотропности среды будем исследовать на примере системы с п = 2 местообитаниями, внутри которых (по-прежнему одинаковым образом) взаимодействуют р — 2 биологических вида с численностями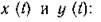
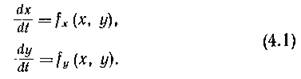
Упрощая обозначения предыдущего параграфа, положим
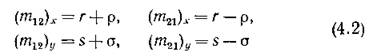
и заметим, что величины р и а характеризуют неодинаковость интенсивностей миграции в противоположных направлениях; случай ,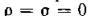 соответствует схемепреды
соответствует схемепреды
дущего параграфа. Пусть для определенности
и ,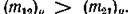 _ _ Тогда ясно, что в принятых
_ _ Тогда ясно, что в принятых
обозначениях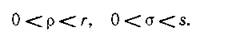
Динамика объединенного сообщества моделируется системой четвертого порядка: 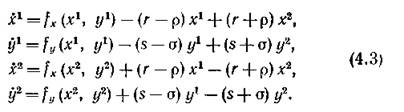
Тот факт, что миграция действительно происходит хотя бы у одного из видов и с неодинаковой по направлению интенсивностью, выражается условиями
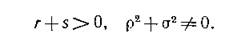
Если в системе (4.3) существует нетривиальное равновесие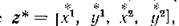 то линеаризация в окрестности
то линеаризация в окрестности
этого равновесия дает матрицу

Действительно, подстановка (4.5) в уравнения стационар-
ного состояния системы (4.3) дает уравнения
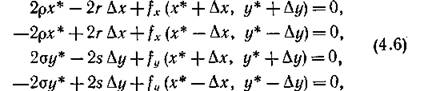
попарное сложение которых и приводит к искомым соотношениям.
Посмотрим, выполняются ли эти условия, например, для вольтерровского описания взаимодействий типа хищник — жеотва без самолимитиоования. т. е. котла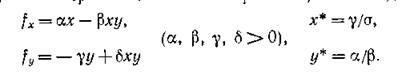
Модели хищник — жертва представляют собой, пожалуй, наиболее интересный объект для исследования эффектов миграции, поскольку здесь, как правило, равновесие имеет нейтрально устойчивый характер, и мы вправе ожидать, что влияние миграции изменит этот характер в ту или иную сторону.
Условия антисимметричности для записанных выше функций требуют, чтобы
Значит, случай (4.7а) возможен либо когда совсем отсутствует миграция хищников, либо когда параметры этой миграции удовлетворяют специальному соотношению, и миграция среди жертв идет преимущественно в том же направлении, что и у хищников (аир совпадают по знаку). При этом сохраняются прежние равновесные значения численностей жертв х*, а численность хищников увеличивается на в той подсистеме, куда миграция предпоч
в той подсистеме, куда миграция предпоч
тительнее (в 1-й в данном случае). Можно сказать, что соотношение (4.7а) между параметрами системы обеспечивает выедание возросшей популяцией хищников избытка жертв, возникающего в 1-й подсистеме за счет предпочтительной миграции. Соотношение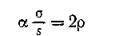
означает, что точка, определяющая систему в пространстве всех параметров модели пппрктипуртся в трехмерное подпространство параметров на часть поверхности
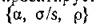
второго порядка (гиперболического параболоида), ограниченную в положительном октанте плоскостью (условие автоматически выполняется).
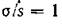
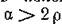
Случай (4.76) возможен, либо когда миграция среди жертв отсутствует совсем, либо когда параметры миграции удовлетворяют специальному соотношению, а миграция хищников идет преимущественно в противоположном направлении (аир противоположны по знаку).
Сохранение равновесных значений у* и перераспределение х* можно интерпретировать так, что избыток хищников, возникающий во 2-й подсистеме благодаря преимущественной миграции, исчезает за счет уменьшившейся (благодаря преимущественной миграции в противоположном направлении) численности жертв в этой подсистеме. Как и в случае (4.7а), точка, изображающая систему в пространстве параметров, проектируется на аналогичную поверхность, но в подпространстве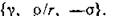
Исследуем устойчивость равновесия в случае объединения двух пар хищник — жертва посредством миграции на уровне жертв с предпочтительным направлением 2->1. Система (4.3) записывается тогда в виде
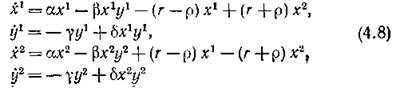

откуда определители Гурвица равны
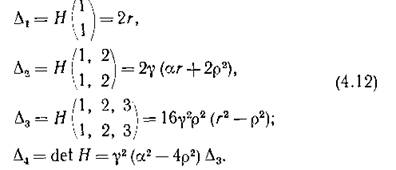
Теорема Рауса — Гурвица гласит, что для отрицательности вещественных частей всех корней действительного
[1] См., например, Гантмахер Ф. Р. Теория матриц. — М., Наука, 1966, с. 488—486,
полинома необходимо и достаточно, чтобы выполнялись условия
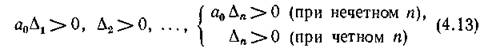
(а0 — коэффициент при старшей степени полинома).
Напомним, что все фигурирующие в формулах (4.12) буквы обозначают положительные параметры, причем г р следует из определения г и р, а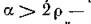 из условия суще
из условия суще
ствования положительной стационарной точки (4.9). Таким образом, за исключением случая, когда р = г, все условия (4.13) выполняются, т. е. для всех собственных чисел матрицы (4.10)
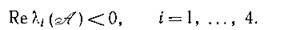
Тем самым, равновесие (4.9) асимптотически устойчиво.
Если вспомнить теперь, что в изолированных парах хищник — жертва равновесие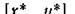 нейтрально устойчиво
нейтрально устойчиво 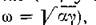
(с частотой колебаний то становится ясно, что
наша система (4.8) представляет собой пример стабилизирующего влияния миграции.
Случай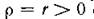 означает, что миграция (с интенсив
означает, что миграция (с интенсив
ностью 2г) идет лишь в одном направлении — из 2-й подсистемы в 1-ю. При этом спектр (корни (4.11)) содержит пару чисто мнимых чисел
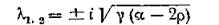
и пару отрицательных чисел
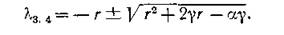
Это значит, что равновесие (4.9) сохраняет в объединенной системе нейтрально устойчивый характер, свойственный изолированным подсистемам, однако частота колебаний в окрестностиоавновесия (в линейном приближении) равняется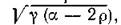 т. е. уменьшается по сравнению
т. е. уменьшается по сравнению
с изолированным случаем. Согласно некоторым экологическим представлениям, меньшая частота колебаний способствует большей стабильности экосистемы, так что, в этом смысле, мы снова имеем пример стабилизирующего вдця- нця миграции.

т. е. однонаправленная миграция хищников увеличивает частоту колебаний в окрестности равновесия по сравнению с изолированным случаем и в этом смысле уменьшает стабильность системы.
Рассмотрим теперь случаи, когда миграция происходит на обоих уровнях, но ее параметры связаны соотношениями (4.7а) или (4.76). Конечно, как отмечал еще В.
Вольтерра, вряд ли можно ожидать, что параметры реальной системы будут связаны какими-либо соотношениями типа равенств, но ради математической полноты мы проведем исследование и этих маловероятных случаев. Когда преимущественное направление миграции у жертв и у хищников одно н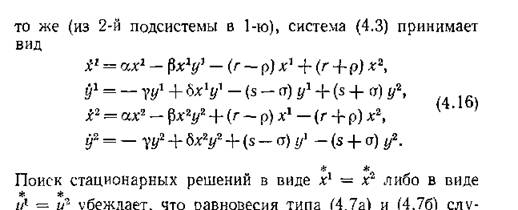
жат единственными стационарными решениями, у которых численности одного уровня в подсистемах одинаковы. (Заметим, что последнее условие может быть проверено в ходе эксперимента.)
Линеаризация (4.16) в точке (4.9) с условием ао — 2ps дает матрицу


Итак, все определители Гурвица положительны, следовательно, по критерию Рауса — Гурвица матрица (4.17) — а вместе с ней и равновесие (4.9) в системе (4.16) — устойчивы.
Равновесие типа (4.76) возможно, когда преимущественные направления миграции на уровне хищников и на уровне жертв противоположны. Чтобы сохранить положительность всех параметров, перепишем систему (4.16) в виде
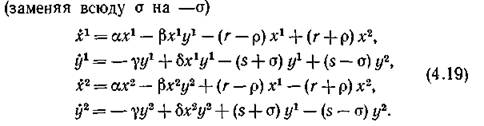
Методом, совершенно аналогичным предыдущему, доказывается, что спектр системы (4.19), линеаризованной в точке
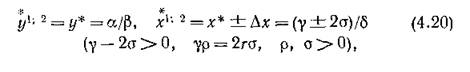
имеет отрицательные действительные части лишь при дополнительном ограничении на множество допустимых зна-
. чений параметров модели
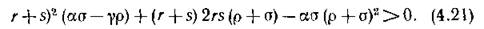
Когда левая часть неравенства .(4.21) обращается в нуль. п гпоитпо ппигитгтотот папа инртп оиык«,1Х Корней
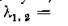
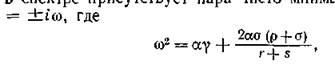
и пара корней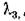 ь с отрицательными действительными частями, так что частота ю колебательных составляющих решений в окрестности равновесия (в линейном приближении) превышает частоту колебаний в изолированной подсистеме.
ь с отрицательными действительными частями, так что частота ю колебательных составляющих решений в окрестности равновесия (в линейном приближении) превышает частоту колебаний в изолированной подсистеме.
Итак, когда миграция, идущая с различной по направлениям интенсивностью на об^их vnnrjHHx, сохраняет одинаковые равновесные значенні в двух подсистемах
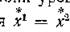
на уровне жертв, она стабилизирует равновесие при всех допустимых значениях параметров модели, а когда сохраняется одинаковая численность на уровне хищников, то стабилизирующее влияние миграции имеет место при дополнительном ограничении (4.21).
В целом же из результатов этого параграфа — при всей частности рассмотренных примеров и ограниченности линейного описания потока миграции — можно сделать вывод, что стабилизирующее влияние миграции на динамику сообществ далеко не однозначно. Напомним, что схемы данного и предшествующего параграфов предполагали также одинаковость уравнений биологических взаимодействий внутри подсистем. Более общая — с этой точки зрения — постановка задачи, предполагающая, однако, достаточную малость параметров миграции по сравнению с параметрами биологических взаимодействий, рассматривается в следующем параграфе.