ПЕРОКСИДНОЕ ОКИСЛЕНИЕ ЛИПИДОВ КАК ОДИН ИЗ КЛЮЧЕВЫХ МЕХАНИЗМОВ МОДИФИКАЦИИ СТРУКТУРНО-ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ БИОМЕМБРАН
Ведущую роль в развитии многих патологических состояний организма играет свободно-радикальное пероксидное окисление липидов (ПОЛ). Наиболее вероятным субстратом ПОЛ в организме являются полиеновые липиды биомембран.
В гомогенных системах процесс ПОЛ протекает по свободнорадикальному цепному механизму. Первичные свободные радикалы появляются в ходе реакции инициирования цепи или катализа, которую считают начальным этапом ПОЛ. Инициирующими факторами ПОЛ в мембранах выступают ферментные системы природных мембранных структур (например, NADPH-зависимая система), ионизирующее и УФ-излучение, различные активные формы кислорода (АФК). Последние способны индуцировать ПОЛ как в темновых процессах (например, при фагоцитозе), так и при воздействии УФ-света, фотоповреждении сетчатки глаза и др.Реакция инициирования цепи имеет вид
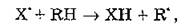
где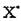 — свободный радикал, инициирующий цепь; RH — ненасыщенная жирная кислота липида; R' — алкильный свободный радикал липида.
— свободный радикал, инициирующий цепь; RH — ненасыщенная жирная кислота липида; R' — алкильный свободный радикал липида.
Затем следуют реакции продолжения цепи:
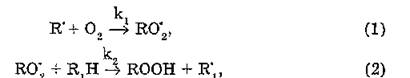
где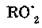 — пероксидный радикал липида; ROOH — гидропероксид липида.
— пероксидный радикал липида; ROOH — гидропероксид липида.
Константа скорости kt реакции составляет 107—108л/моль- с, а энергия активации практически равна нулю. Следовательно, при обычных концентрациях кислорода (>10~п моль/л) все радикалы R' превращаются в R0'2. Реакция (2) также имеет низкую энергию активации и высокую константу скорости к2, величина которой зависит от типа окисляющегося соединения.
Разветвление цепи осуществляется в соответствии с реакцией
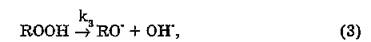
где RO' — алкоксильный радикал; ОН' — гидроксильный радикал.
С реакциями продолжения конкурируют реакции, приводящие к обрыву цепи. Возможны следующие варианты спонтанного обрыва цепи: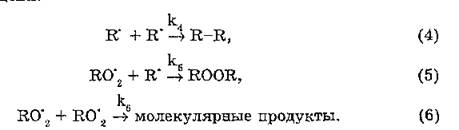
Сущность цепного процесса окисления состоит в чередовании двух реакций — образования иероксидного радикала липида R0'2, а также гидропероксида ROOH и нового радикала липида R': 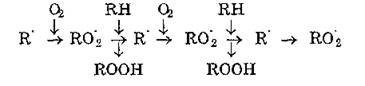
Таким образом, в процесс вовлекаются все новые молекулы липида (RH) и кислорода, при этом накапливаются гидропероксиды, а число радикалов R' и R0‘2 не изменяется в соответствии с принципом неуничтожимое™ свободной валентности.
Генерирование липопероксидов в организме сопряжено с нормальными метаболическими реакциями, осуществляемыми специализированными ферментными системами: NADPH-зависимыми микросомальными оксигеназами, циклооксигеназами и липоокси- геназами, и служит источником биосинтеза внутриклеточных медиаторов — простагландинов, тромбоксанов, простациклина, лей- котриенов и липоксинов. Свободная полиненасыщенная жирная кислота, в частности арахидоновая, образовавшаяся в результате ферментативного гидролиза (З-ацилов лецитинов фосфолипазой А2, может окисляться по двум альтернативным путям — цикло- или липооксигеназному. При циклооксигеназном окислении арахидо- ната образуются циклические зндопероксиды — простагландины G-2 и Н2, служащие универсальными метаболическими предшественниками различных простагландинов (PGE2, PGE2a, PGD2), тромбоксанов (ТХА2, ТХВ2) и простациклина (PGI2). Продуктами липооксигеназного окисления арахидоната являются алифатические гидропероксиды — гидропероксиэйкозатетраеновые кислоты (НРЕТЕ) — промежуточные продукты биосинтеза нового класса биологически активных веществ — лейкотриенов (LTA4, LTB4, LTC4, LTD4) — рис.
27. Производные С20-полиненасыщенных жирных кислот типа арахидоновой, так называемые эйкозаноиды, участвуют в осуществлении защитных реакций клеток желудка, сердца и других органов от повреждений, способствуют развитию воспаления, стимулируют сокращение гладкой мускулатуры. Циклооксигеназные системы обнаружены в большинстве тканей животных, липооксигеназные — в различных животных клетках и тканях: лейкоцитах, тромбоцитах, ретикулоцитах; легких, селезенке, семенниках. Между липооксигеназным и циклооксигеназным путями окисления полиненасыщенных жирных кислот в организме существует тесная взаимосвязь. Ингибирование циклооксигеназного пути окисления этих клеток in vivo сопровождается активацией их липооксигеназного окисления, т.е. липооксигеназный путь окисления может использоваться для быстрой утилизации избыточного субстрата биосинтеза простагландинов.Регуляция процессов ПОЛ в организме также осуществляется ферментативным путем. В микросомах наряду с фермента-
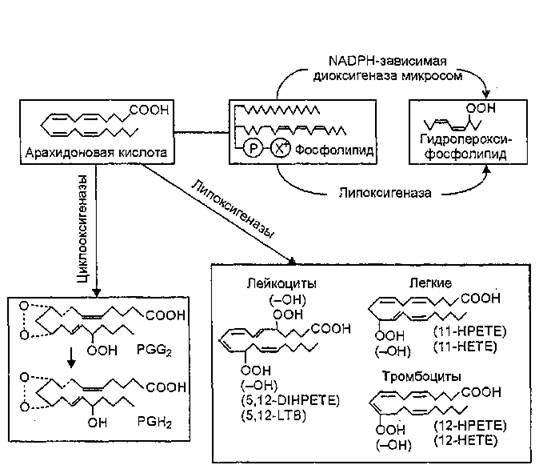
Рис. 27. Основные пути ферментативного биосинтеза пероксидов липидов в тканях млекопитающих (В. 3. Ланкин, 1984): НЕТЕ — гид- роксикозатетраеновые кислоты; НРЕТЕ — гидропероксиэйкозатетра- еновые кислоты; DIHPETE — дигидропероксиэйкозатетраеновая кислота; LTB — лейкотриены; PGG2, PGH2 — простагландини
тивным синтезом пероксидов липидов происходит и их ферментативная утилизация, так как терминальная оксидаза цепи переноса электронов в микросомах — цитохром Р450 — может функционировать как активная пероксидаза. В цитозоле клеток содержатся восстанавливающие пероксиды липидов глутатионпер- оксидазные и глутатиои-Э-трансферазиые системы, одной из функций которых является ограничение распространения неферментативных реакций ПОЛ в случае “утечки” свободнорадикальных продуктов ферментативного ПОЛ. Существование специализированных ферментных систем синтеза и утилизации активных форм кислорода и пероксидов липидов делает маловероятным (если не невозможным) протекание в нормальных тканях животных неспецифического процесса — неферментативного ПОЛ с физиологически значимой интенсивностью.
Необходимо отметить, что в клетке существует несколько пусковых и защитных систем, которые рассматриваются как факторы, влияющие на скорость окисления липидов на разных стадиях (зарождение и продолжение цепи; разветвление; обрыв цепи):
— системы, ответственные за строго определенную структурную организацию липидов и влияющие на скорость окисления липидов на разных стадиях (зарождение и продолжение цепи; разветвление; обрыв цепи);
— системы ферментов, ответственных за образование и гибель активных форм кислорода и свободных радикалов или участвующих в разложении пероксидов нерадикальным путем;
— системы, регулирующие обмен фосфолипидов мембран и влияющие на скорость инициирования и продолжения цепей путем изменения состава ненасыщенных жирных кислот фосфолипидов;
— системы низкомолекулярных регуляторов, выполняющих роль инициаторов, ингибиторов и влияющих на стадию инициирования, разветвления и обрыва цепи.
ПОЛ представляет собой один из важнейших универсальных процессов повреждения мембранных систем, изменяющий химический состав, физические параметры, ультраструктурную организацию и функциональные характеристики биомембран. ПОЛ вызывает обновление липидного состава мембран вследствие удаления легко окисляющихся липидов — фосфатидилсерина, фосфати- дилэтаноламина, фосфатидилинозитола. При ПОЛ возрастает скорость процессов “флип-флоп”-переходов. ПОЛ приводит к увеличению вязкости мембран в результате уменьшения содержания жидких липцдов в бислойных участках, появления поперечных межмолекулярных сшивок и возрастания доли упорядоченных липидов с ограниченной подвижностью. Отрицательный заряд на поверхности мембран увеличивается, что обусловлено вторичными продуктами ПОЛ (эпоксиды, кетоны, малоновый диальдегид и др.), содержащими карбонильные и карбоксильные группы. Мембраны эритроцитов, митохондрий, саркоплазматического ретикулума, лизосом становятся проницаемыми для различных ионов, неэлектролитов, макромолекул. Изменяются свойства мембранных белков: Саг+-АТФазы, Na+, К+- АТФазы, родопсина, фосфолипазы. Эти функциональные проявления ПОЛ определяют формирование многих патологических состояний организма, возникающих при неблагоприятных условиях и повреждающих воздействиях.
Как уже было отмечено выше, в качестве инициаторов ПОЛ выступают активные формы кислорода или активные кислородные метаболиты. Рассмотрим более подробно механизмы их образования и пути утилизации, функциональное значение и роль в развитии патологических состояний организма человека.
3.2.
Еще по теме ПЕРОКСИДНОЕ ОКИСЛЕНИЕ ЛИПИДОВ КАК ОДИН ИЗ КЛЮЧЕВЫХ МЕХАНИЗМОВ МОДИФИКАЦИИ СТРУКТУРНО-ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ БИОМЕМБРАН:
- Структурно-функциональные модификации молекулярных компонентов биомембран под действием УФ-излучения
- 7. Концепция А.Р. Лурии о трех основных структурно-функциональных блоках. 8. Специфика вклада структурно-функциональных блоков в осуществление высших психических функций.
- Радиационно-химические превращения структурных компонентов биомембран
- § 2. Концепт «Бог» как один из ключевых концептов русской наивной картины мира
- Публицистический стиль как функциональная разновидность современного русского литературного языка. Структурный принцип организации публицистического стиля.
- Методы модификации психических состояний объекта
- Психология стресса и функциональных состояний работника
- Нарушение структурно-функциональных свойств клеточных мембран
- 11. Характеристика 2-го структурно-функционального блока мозга.
- 43. Управление функциональным состоянием предпринимателя
- Структурные нарушения и функциональные расстройства мозга
- 12. Характеристика 3-го структурно-функционального блока мозга.
- 10. Характеристика 1-го структурно-функционального блока мозга.
- 27. Третий структурно – функциональный блок мозга. Агранулярная и гранулярная кора.
- 7.1. Равновесное состояние изолированной системы как состояние с максимальной энтропией
- 3. Какова функциональная структура механизмов перцепции?
- А. Мартине ОСНОВЫ ОБЩЕЙ ЛИНГВИСТИКИ функционально-структурные основы ЛИНГВИСТИЧЕСКОГО ОПИСАНИЯ
- Тест дифференциальной самооценки функционального состояния (САН)
- Механизмы регулирования функциональной активности ферментов и ферментных систем в клетке