Структурно-функциональные модификации молекулярных компонентов биомембран под действием УФ-излучения
Биологически значимое УФ-излучение занимает участок спектра от 200 до 400 нм и подразделяется на три области: коротковолновую (200—280 нм), средневолновую (280—315 нм) и длинноволновую (315—400 нм).
Поверхности Земли достигает УФ-свет в диапазоне длин волн 287—400 нм с энергией квантов ~3—4 эВ. Коротковолновая часть спектра УФ-излучения Солнца полностью поглощается озоновым слоем атмосферы.В биосистемах УФ-свет индуцирует главным образом фото- деструктивные реакции, связанные с фотохимическими превращениями белков и нуклеиновых кислот, относящихся к основным акцепторам УФ-излучения в клетке. Однако в биомембранах и других липидных системах под действием УФ-излучения эффективно протекает процесс пероксидного фотоокисления (ПФО) ненасыщенных жирных кислот липидов, который приводит к значительным изменениям структурно-функционального состояния всех мембранных компонентов. Таким образом, биомембраны содержат различные хромофорные группы, поглощающие энергию УФ-излучения в разных диапазонах длин волн, — ароматические и серосодержащие аминокислоты мембраносвязанных белков, полиненасыщенные жирные кислоты липидов, а также коферменты, включающие пиридиннуклеотиды, флавины, кофермент Q, железопорфирины, витамины.
Результаты исследования фотохимического действия оптического излучения на биомембраны клеток показывают, что наиболее эффективным является УФ-излучение с длинами волн КО-
роче 300 нм. Главный путь фотолиза мембранных липидов - фотоокисление цепей полиненасыщенных жирных кислот фосфолипидов. Следует отметить, что липиды поглощают УФ-свет в коротковолновой области спектра (Х< 240 нм), а максимумы поглощения ненасыщенных жирных кислот находятся в области < 220 нм. Однако фотоокисление липидов развивается при воздействии на мембраны более длинноволнового излучения, что обусловлено поглощением УФ-света гидропероксидами липидов, присутствующими в норме в липидных системах вследствие протекания ферментативных процессов ПОЛ (см.
главу 3). Диеновые гидропероксиды жирных кислот имеют максимум светопогло- щения при - 233 нм, а триеновые гидропероксиды — при - 270 нм. Процесс ПФО липидов можно представить в общей форме в виде совокупности элементарных реакций:
и др. — константы скоростей реакций. Реакции (а) и (е) — фотохимические, реакции (б)—(д) — темновые. Реакция (а) носит название реакции фотоинициирования, если ROOH — предсуще-
ствующие гидропероксиды, или разветвление цепей окисления, (б), (в), (г), (д) — соответственно продолжение цепи, “квадратичный” обрыв цепи (диспропорционирование радикалов ROO'), “линейный” обрыв цепи на антиоксиданте, инициирование цепи окисления свободным радикалом антиоксиданта. УФ-облучение мембран и других липидных систем индуцирует темновое пероксид- ное окисление ненасыщенных жирных кислот липидов. Процесс автоокисления в облученных мембранах протекает гораздо медленнее фотоокисления, но за длительный темновой период он может приводить даже к более сильному окислению мембранных липидов, чем фотоокисление. Темновое автоокисление липидов в облученных мембранах блокируется антиоксидантами в низких концентрациях. Под действием УФ-излучения в некоторых клетках стимулируется темновое ферментативное пероксид- ное окисление липидов с участием циклооксигеназы, в результате которого образуются простагландины. Это явление обнаружено в коже и играет важную роль в развитии эритемной реакции в ответ на УФ-облучение.
Следует подчеркнуть, что процесс пероксидного фотоокисления липидов (ПФОЛ) является двухквантовым и протекает по сложному механизму. Первая фотохимическая стадия его — генерация гидропероксидов в результате присоединения кислорода; вторая стадия, не зависящая от присутствия кислорода, — фотолиз гидропероксидов до вторичных продуктов (альдегидов и кетонов), поглощающих УФ-излучение в диапазоне длин волн 260—280 нм. При воздействии видимого света и длинноволнового УФ-излучения процесс заканчивается в основном на стадии образования гидропероксидов, а при облучении липидных систем более коротковолновым светом (X - 233 нм) накапливается большое количество вторичных продуктов.
Один из вторичных продуктов ПОЛ — малоновый диальдегид — при взаимодействии с тиобарбитуровой кислотой (ТБК) дает окрашенное соединение, поглощающее свет с длиной волны -533 нм. Эту реакцию используют для определения уровня интенсивности протекания ПФОЛ. Второй подход, позволяющий судить об эффективности пероксидного окисления липидов, связан с регистрацией хемилюминесценции, являющейся следствием реакции диспропорционирования пероксидных радикалов липидов.
Каково же значение пероксидного окисления липидов в фотоповреждении биологических мембран? Фотолиз липидов мо
жет приводить прежде всего к существенным нарушениям структурной организации компонентов биомембран. Продукты фотоокисления липидов обладают достаточно выраженными токсическими свойствами. Пероксиды липидов и продукты их дальнейших превращений (альдегиды и кетоны) могут индуцировать повреждение белковых молекул, в частности, их сульфгидриль- ных групп. Повреждение белков может быть обусловлено как их окислением, так и образованием стабильных ковалентных связей между молекулами белков и продуктами пероксидного фотоокисления липидов. Последние способны инактивировать многие мембраносвязанные ферменты. Кроме того, вышеуказанные продукты окисляют также такие биологически важные соединения, как цистеин, глутатион, нуклеотиды, витамины А и Д, липо- евую кислоту и др.
Таким образом, действие УФ-излучения приводит к значительным модификациям структурно-функционального состояния компонентов различных мембранных структур и их взаимодействий: изменяется пассивная проницаемость липидного бислоя для ионов, разобщается окислительное фосфорилирование, изменяются конформация и каталитическая активность мембраносвязанных ферментов, нарушаются межклеточные взаимодействия, происходит набухание и лизис клеток и их органелл.
Ряд исследователей считает, что мембранные эффекты УФ- облучения в основном индуцированы ПФОЛ и лишь частично обусловлены фотохимическими превращениями белков.
Напомним, что происходящие в белках под воздействием УФ-излучения нарушения их структурно-функционального состояния индуцируются в основном поглощением энергии УФ-света остатками ароматических аминокислот: триптофана, тирозина, фенилаланина, а также остатками серосодержащих аминокислот цистеина и цистина. Под влиянием УФ-света происходит фотоионизация остатков аминокислот с образованием сольватирован- ного электрона и катион-радикала аминокислоты: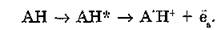
Катион-радикал — сильная кислота и при 77—140 К диссоциирует на протон и нейтральный радикал:
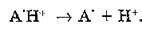
Нейтральные радикалы являются неустойчивыми соединениями и при температуре выше 200 К вступают в дальнейшие пре-
вращения. Взаимодействуя с соседними группами полипептид- ной цепи, они могут образовывать межмолекулярную сшивку, являющуюся стабильным фотопродуктом:
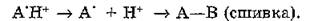
В присутствии кислорода происходит фотоокисление триптофана, тирозина и фенилаланина. Большое значение в фотохимии белков имеют фотосенсибилизированные реакции с участием сольватированного электрона. Наибольшим сродством к электрону обладают цистин и цистеин. Они быстро разрушаются в результате взаимодействия с сольватированными электронами е~, выбитыми из ароматического кольца:
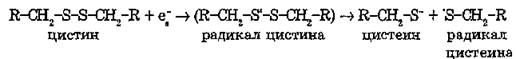
Если фотолизу подвергается остаток аминокислоты, непосредственно входящей в состав активного центра фермента, уже этого может быть достаточно, чтобы белок потерял ферментативную активность. Если сшивка находится вне активного центра фермента, то она способна изменить баланс водородных, гидрофобных и других слабых связей (множественные разрывы связей), поддерживающих нативную конформацию макромолекулы.
В большинстве случаев конечным результатом действия УФ-света на белки является их инактивация, т.е. потеря ферментативной, регуляторной, транспортной, гормональной, иммунологической и других видов активности. Вместе с тем для ряда белков (карбоксипептидаза А, супероксиддисмутаза, цитохром С, гемоглобин) выявлена их активация под влиянием УФ-излучения.Необходимо отметить, что фоточувствительность мембраносвязанного белка может существенно отличаться от таковой для этого же биополимера в свободном состоянии (в растворе), что обусловлено особенностями микроокружения белковых молекул в мембране, их взаимодействием с другими компонентами надмолекулярного комплекса.
Иными словами, уровень фоточувствительности мембранных белков-ферментов будет эффективно контролироваться структурным состоянием их микроокружения.
Доказательства мембранного контроля УФ-чувствительности ферментов были получены С. В. Коневым и И. Д. Болотовским (1979) на основании анализа изменений величины поперечного
сечения инактивации ацетилхолинэстеразы в свободном состоянии, в составе интактной мембраны, в мембране, обработанной фосфолипазами А2, С и D, в мембране, обедненной по холестерину, Под поперечным сечением инактивации (о) понимают произведение поперечного сечения поглощения (S) активного света и квантового выхода инактивации („/(Фі)СІ = = 1 + кД),ЯЪ
где