Сравнение химической и электрической проводимости
Не менее 80% известных фармакодинамических агентов действуют исключительно на нерв или нервно-мышечное соединение. Большинству нервных клеток присущи определенные структурные особенности, позволяющие выделить три основные обла-
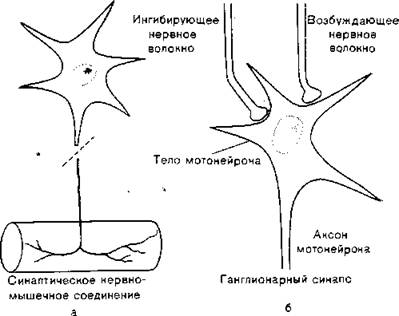
Рис.
7.1. Соединение, образованное: а) нервом и мышцей, б) тремя нервами.сти клетки: клеточное тело, дендриты и аксон. Обычно тело нейрона имеет сферическую или пирамидальную форму. Аксон отличается от дендритов не только строением, но и свойствами наружной мембраны и, как правило, длиннее и тоньше дендритов. В области синапса аксон расширяется, образуя на конце пресинаптическую бляшку, которая представляет собой передающую информацию часть контакта. Постсинаптическая мембрана, на которой расположены рецепторы, отделена от бляшки синаптической щелью, ширина которой обычно 20—40 нм. Скопление нейронов и нервных волокон могут образовывать нервные узлы — ганглии. На рис. 7.1 схематично представлены синапс нерв — нерв и нервно-мышечный синапс, в сущности подобные друг другу [Eccles, 1965; Katz, 1966].
В среде, окружающей нейрон, концентрация ионов натрия значительно выше, чем во внутренней, тогда как во внутренней среде значительно выше концентрация ионов калия. Благодаря различию концентраций ионов по разные стороны цитоплазматической мембраны в нервных и мышечных клетках обычно возникает отрицательный потенциал («потенциал покоя») величиной от 50 до 100 мВ (по сравнению с внешним, принимаемым за 0). Таким образом, мембрана представляет собой как бы миниатюрную батарею с отрицательным полюсом внутри. При снижении потенциала говорят, что мембрана деполяризуется. Через такую липопротеидную мембрану возможен активный транспорт в обоих направлениях (разд. 3.2). Транспорт ионов натрия из клетки и ионов калия внутрь ее, в направлении, противоположном электрохимическому градиенту этих ионов, осуществляет внутренний мембранный белок — Ыа+/К+-АТФаза.
Распространение нервного импульса по нервному волокну —
сложный физико-химический процесс деполяризации мембраны нервного волокна (в самой нервной системе — для двигательного волокна либо в периферических органах — для сенсорного волокна). При возникновении нервного импульса (например, в основании аксона) трансмембранная разность потенциалов в этом месте локально понижается, при этом на короткое время повышается местная проницаемость мембраны для ионов натрия непосредственно перед областью с изменившимся потенциалом. Ионы натрия быстро входят внутрь нервного волокна, изменяя в этом участке отрицательный внутренний потенциал мембраны на положительный. Это приводит к закрыванию натриевых каналов и открыванию калиевых, что позволяет ионам калия выходить наружу. Поток ионов восстанавливает потенциал внутри волокна до величины потенциала покоя. Все эти изменения происходят на небольшом участке нервного волокна, но вызванные ими потоки ионов деполяризуют следующий участок мембраны, и так далее. В результате нервный импульс, являющийся по существу кратковременным изменением мембранного потенциала («потенциала действия»), распространяется по волокну со скоростью, зависящей от диаметра волокна. Обычно скорость распространения нервного импульса изменяется в пределах от 0,1 до 100 м/сек-1 [Hodgkin, 1964].
Позднее было показано, что катионные каналы гигантского аксона кальмара могут блокироваться ионами водорода. Исходя из величин напряжения, требуемого для преодоления этого блока, было рассчитано, что прохождение катиона натрия по натриевым каналам контролируется двумя кислотами с рКа 4,6 и 5,8 соответственно [Wanke, Carbone, Testa, 1980].
Быстрый натриевый ток избирательно блокируется тетродо- токсином (7.46)—сферической молекулой пергидрохиназолина, содержащей высокоосновную гуанидиновую группу, выступающую из нее наподобие языка. В природе это соединение встречается в некоторых видах рыб, амфибий и моллюсков. Перекрывая натриевый ток, тетродотоксин блокирует генерирование потенциала действия, а следовательно, и его передачу.
Так как передача импульса в синапсах при этом не нарушается, млекопитающие обычно погибают от остановки дыхания [Narahashi, Moore, Scott, 1964]. На транспорт ионов калия тетродотоксин не влияет.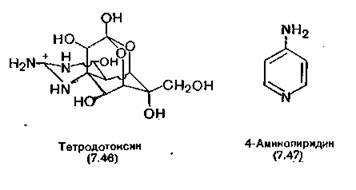
Гуанидиний — один из немногих катионов, способных, подобно иону натрия, вызывать потенциал действия [Watanabe et al., 1967], поэтому логично предположить, что гуанидиновая перегруппировка тетродотоксина входит в натриевый канал, а остальная часть молекулы его закрывает. Сакситоксин — пергидропурин с двумя гуанидиновыми группировками (выделен из океанских одноклеточных динофлаггелятов) действует почти идентично тетродотоксину [Gage, 1971; Goto et al., 1965].
Более сильное гетероциклическое основание 4-аминопиридин (7.47) избирательно блокирует калиевые каналы, увеличивая тем самым приток ионов кальция при деполяризации нервных окончаний [Thesleff, 1980]. В последнее десятилетие в медицинскую практику вошли блокаторы кальциевых каналов в качестве сосудорасширяющих средств, например верапамил, нифеди- пин (разд. 14.2).
Говоря о нормальном возбуждении нервного окончания, следует заметить, что прохождение одиночного импульса вызывает лишь небольшое повышение внутриклеточного соотношения Na+:K+, в конечном счете восстанавливающегося до нормального (без изменения потенциала покоя) в результате транспорта ионов, требующего затраты энергии — так называемого