Клеточная стейка
Наличие клеточной стенки — это особое свойство растений {включая грибы), отличающее их от других живых организмов.’ Клеточная стенка обеспечивает клеткам большую прочность, но' не регулирует проницаемость, что является функцией совершен-? но другой структуры — плазматической мембраны (см.
ниже). У всех представителей животного царства клеточные стенки полностью отсутствуют. Большинство одноклеточных организ-і мов, включая Amoeba и Trypanosoma, также лишены этой стен-, ки. Лишь некоторые из них окружены стенками из белка) (Eimeria) или целлюлозы (Chlamydomonas). |А. Клеточные стенки многоклеточных растений состоят ИЭ| микрофибрилл целлюлозы различной длины, диаметром 10—|
20 нм, включенных в аморфный матрикс из гемицеллюлозы и пектинов, представляющих собой частично О-метилированную кальциевую соль полимеров галактуроновой кислоты. Клеточную стенку синтезируют ферменты плазматической мембраны [Siegel, 1962].
Б. Клеточная стенка грибов представляет собой мозаику из различных углеводов с отдельными включениями липидов и белков. Основным углеводом является хитин (поли-1Ч-ацетилглюко- замин), хотя у некоторых видов дрожжей хитин отсутствует. Хитин составляет основную часть внешнего покрова насекомых и ракообразных.
Дрожжи — это одноклеточные грибы; их клеточные стенки состоят из двух тесно переплетенных структур, причем для сохранения характерной формы клеток достаточно наличия любой из них. Одна из этих структур состоит целиком из глюкана [полиангидрид глюкозы), вторая представляет собой маннан- протеиновый комплекс, компоненты которого соединены между собой дисульфидными связями. Только разрушение обеих структур приводит к выходу цитоплазмы в окружающую среду [Bacon et at, 1965].
В отличие от клеток высших организмов клетки грибов испытывают высокое внутреннее осмотическое давление, поэтому если разрушить клеточную стенку, например, действием хитина- зы (выделенной из улиток), то клетки лопаются; этого можно избежать, если повысить осмотическое давление снаружи.
Механизм действия фунгицида пентахлорнитробензола (квинтозе- на) основан на его способности снижать содержание хитина в стенках мицелия [Macris Georgopoulis, 1969].В. В бактериальных клетках также существует высокое осмотическое давление, особенно у грамположительных микроорганизмов [Brown, 1964; Mitchell, Moyle, 1957]. От разрыва их удерживает толстая стенка, на долю которой приходится до 25% сухого веса клетки. В процессе роста протопласта (т. е. всего содержимого клетки, помимо стенки) должен происходить и синтез материала клеточной стенки. Механизм действия некоторых антибиотиков, применяемых в медицине, заключается в нарушении синтеза клеточной стенки (см. ниже). Поэтому при росте бактерий в результате ослабления клеточной стенки наступает разрыв бактериальных клеток под действием собственного осмотического давления. Впервые это явление наблюдал Leder- berg (1957) на примере пенициллина. Он отметил, что лизис можно предотвратить, используя культуральные среды с высоким содержанием солей.
Величина эффективного диаметра пор в стенке бактериальных клеток составляет приблизительно 1 нм [Mitchell, Moyle, 1956]. Стенка грамположительных бактерий толщиной 15— 50 нм примерно наполовину состоит из муреина [так называл эту молекулу Park (1966), первый исследователь молекулярного механизма действия пенициллина] — полисахаридно-полипеп-
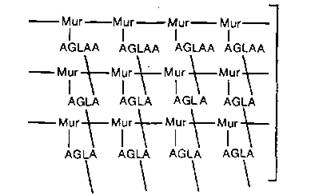
Рис. 5.3. Фрагмент муреина: поперечная сшивка единиц муропептида соответствующими аминокислотами. Mur — дисахарид, AGLA — четыре аминокислоты, AGLAA — пять аминокислот.
тидного полимера. Примерно 25 лежащих друг на друге слоев муреина придают стенке прочность. Остальные компоненты стенки бактерий представлены в основном тейхоевой кислотой (см. ниже). Муреин является специфическим субстратом для лизоцима, противобактериального фермента, содержащегося в слезах и других секретах организма, а также в белке яйца. Лизоцим используют фаги вирусов для проникновения в бактериальную клетку.
Стенка грамотрицательных бактерий устроена значительно сложнее. Под электронным микроскопом отчетливо видны концентрические слои. Уникальным свойством грамотрицательных бактерий является то, что в них муреин с наружной стороны покрыт второй полупроницаемой мембраной, состоящей из липопротеидов и липополисахаридов. В стенках этих бактерий отсутствует тейхоевая кислота, а муреин составляет всего лишь 5—20% всей массы стенки. Его слои, толщиной около 2 нм, с внутренней стороны тесно связаны с плазматической мембраной, их целостность зависит от наличия магния и кальция. Предполагают, что наружная мембрана возникла в результате естественного отбора для защиты муреина от разрушения лизоцимом. Это подтверждается и тем фактом, что муреин грамотрицательных организмов значительно более однороден, чем муреин грамположительных бактерий, для которых естественный отбор был направлен на повышение устойчивости муреина к внешним воздействиям [Nikaido, 1979]. Эта структура грамотрицательных бактерий создает трудности при разработке лекарственных веществ, после преодоления которых уже не приходится сталкиваться с таким количеством различий, как у грамположительных организмов.
Муреин (известный также под названием пептидогликан) представляет собой поперечно-сшитый полимер неопределенного размера. Весьма вероятно, что всего лишь одна молекула муреина обвалакивает всю бактерию. Когда в процессе роста клетки возникает необходимость увеличить размер молекулы
муреина, происходит встраивание дополнительных частей в разрезы, образующиеся под действием специальных цитоплазматических ферментов. Молекула муреина состоит из линейных цепей дисахаридов, поперечно-сшитых боковыми цепями аминокислотных остатков (рис. 5.3).
Г. Биосинтез муреина. Его первой стадией является синтез характеристичного моносахарида. Вначале в цитоплазме из N-ацетилглюкозамин-І-фосфата и УТФ образуется уридин-N- ацетилглюкозамин. Затем в положение 3 этого нуклеотида (в две стадии) присоединяется остаток молочной кислоты и образуется уридиндифосфо-Мі-ацетилмураминовая кислота [аце- тилмураминовая кислота — это З-О-О-лактил-М-ацетилглюкоза- мин (5.9), сахар, встречающийся только в клеточной стенке прокариот].
К ней присоединяются пять аминокислотных остатков, причем концевой всегда является пара-О-аланил-О-аланин. В зависимости от вида состав этого пентапептида может меняться, например, последовательность для Staphylococcus aureus представлена формулой (5.10).
Затем этот моносахарид превращается в дисахарид. На первой стадии он ковалентно присоединяется к изопреноидному спирту С55 плазматической мембраны, отщепляя при этом УМФ [Higashi, Strominger, Sweeley, 1967]і. После этого молекула N-ацетилглюкозамина реагирует с остатком лизина, с которым соединены и пять остатков глицина (образование этой цепи 215
идет без участия рибосом и в направлении, противоположном нормальному пептидному синтезу). Полученный таким образом продукт обычно называют «дисахарид декапептида».
Следующей стадией является полимеризация. Разрывается связь дисахарида декапептида с Css-компонентом мембраны и образуется связь, соединяющая положение 1 остатка N-ацетил- мураминовой кислоты с гидроксильной группой в положении 4 концевого остатка N-ацетилглюкозамина другого дисахарида. Повторение этого процесса (примерно 50 раз) приводит к росту полисахаридной цепи [Strominger et al., 1967].
Последняя стадия — образование поперечных сшивок в результате реакции транспептидации. Эта реакция не требует притока внешней энергии и протекает между концевой аминогруппой пентаглициновой боковой цепи и карбонильным остатком предпоследнего остатка D-аланина в другой пентапептид- ной цепи. При этом отщепляется одна молекула D-аланина и образуется новая пептидная связь [Wise, Park, 1965: Tipper, Strominger, 1965]. Вследствие повторения этого процесса образуется слой муреина.
Несколько иначе устроен муреин грамотрицательных организмов. В нем отсутствует пентаглициновая цепь, а сшивка образуется между концевой аминогруппой мезо-диаминопимели- новой кислоты (занимающей место лизина) и предпоследним D-аланином.
Диаминопимелиновая кислота также ковалентно связана с липопротеином [Hofschneider, Martin, 1968]. Несмотря на малое содержание, именно муреин определяет прочность стенки грамотрицательных бактерий. Его удаление из клеточной стенки приводит к разрыву клетки [Mandelstam, 1962].Тейхоевые кислоты — это полимеры глицерофосфата и ри- битфосфата, расположенные на С55-стороне плазматической мембраны грамотрицательных бактерий. К этому полимерному остову, в зависимости от вида бактерий, присоединены различные заместители (D-аланин, глюкоза, галактоза или аминосахара). Тейхоевые кислоты принимают участие в обмене ионов между клеточной стенкой и плазматической мембраной, накапливают ионы магния и определяют групповые антигенные свойства клеточной стенки. Больше нигде в природе не встречаются [Baddiley, Hancock, Sherwood, 1973].
Стенки спор бактерий построены так же, как и клеточные стенки, но, кроме этого, покрыты еще комплексом кальция с дипиколинатом, в свою очередь укрытого снаружи белковой оболочкой с множественными дисульфидными связями [Gould, Hitchins, 1963].
О строении бактериальных клеточных стенок см. Gale и сотр. (1981) и Franklin, Snow (1981).
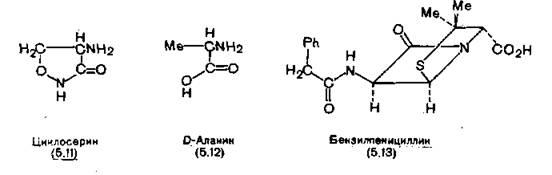
Механизм действия некоторых антибиотиков заключается в нарушении различных стадий биосинтеза клеточной стенки, бактерий. Структурный аналог D-аланина циклосерин (5.11), применяющийся при' устойчивых формах туберкулеза (разд. 2Ї6
9.4.3), ингибирует два фермента: один из них рацемизует L-аланин в D-аланин (5.12), а второй синтезирует из D-аланина О-аланил-О-аланин [Strominger, Threnn, Scott, 1959]. Со вторым ферментом циклосерин связывается в 100 раз эффективнее, чем нормальный субстрат. К аналогам-антагонистам аланина, имеющим значение для клинической практики, относятся 3-фтор- D-аланин и его 2-дейтеропроизводное [Kollonitsch et al., 1973] и алафосфалин (3.50) (разд. 3.6).
Механизм противомикробного действия бензилпенициллина (5.13), первого представителя пенициллинов, заключается в образовании ковалентной связи между ним и «пептидогликан- транспептидазой», ферментом, в норме образующим поперечные сшивки в муреине на последней стадии биосинтеза [Izaki, Mat- suhashi, Strominger, 1968]. В результате этого растущая бактерия теряет способность строить новую стенку и погибает (см. также разд. 13.1 и 13.2).