4.0.6. Лекарственные вещества — ингибиторы синтеза РНК
Родоначальники антибиотиков ряда рифамицина были выделены из Streptomyces mediterranei. Избирательность действия этих соединений можно повысить химическими модификациями, и поэтому вместо природного рифамицина (3.47, а) в основном применяют его производное рифампицин (3.47,6).
В сочетании с изониазидом (разд. 11.9) он широко используется при некоторых формах туберкулеза, но его применение ограничено высокой стоимостью препарата [American Thoracic Society, 1980]. Рифампицин является наиболее избирательным из всех известных противобактериальных препаратов и может с успехом применяться при многих инфекционных заболеваниях, но обычно его назначают при стафилококковых инфекциях, устойчивых к пе- .нициллину.По химическому строению рифамицины относятся к группе
ІІ60
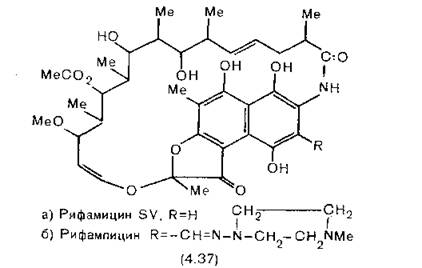
природных соединений, называемых ансамицинами (от латинского ansa — ручка). Название анса-соединения было введено Liittringhaus и Gralheer для соединений, содержащих ароматическое кольцо (в данном случае нафталиновое), с огибающей его «ручкой», образованной присоединенной к циклу в двух местах алифатической цепью [Maggi et al., 1966, 1968]. Для того чтобы такие соединения обладали биологическим действием, необходимо наличие по меньшей мере трех гидроксильных групп в их молекуле. Синтетические модификации рифамицина сводятся преимущественно к введению заместителя в положение 3. Восстановление двойных связей в алифатическом цикле снижает активность пропорционально уменьшению жесткости молекулы.
Рифамицины взаимодействуют с р-субъединицей бактериальной ДНК-зависимой РНК-полимеразы с образованием очень прочного нековалентносвязанного комплекса с соотношением компонентов 1 : 1, в результате чего прекращается только синтез РНК, но не ДНК и белков. В случае рифамицин-устойчивых организмов такой комплекс не образуется.
ДНК-зависимая РНК- полимераза млекопитающих не имеет ^-субъединицы, и поэтому избирательность действия рифамицинов так высока [Tocchini- Valenti, Marino, Colvill, 1968].В природе существуют и другие ансамицины, например стреп- товарицин, выделяемый из другого вида Sterptomyces, действие которого аналогично действию рифамицина, ио менее избирательно; аманитин (из гриба Amanita phalloides), в отличие от рифамицина действующий только на ДНК-зависимую РНК-полимеразу эукариот и не влияющий на этот фермент у прокариот; и маутансин (из коры африканского цветкового растения), противоопухолевые свойства которого клинически не исследованы.
Дактиномицин (4.38), антибиотик ярко-красного цвета, выделенный Waksman и Woodruff в 1940 г., является производным аминофеноксазина и содержит две одинаковые циклические боковые цепи с одной эфирной и пятью пептидными связями (4.39). В каждую боковую цепь входят остатки N-метилвалина, саркозина, пролина, валина и треонина [Brockmann, 1960]. По данным ЯМР-спектров —NH валина образует сильную водородную связь с СО саркозина.
Дактиномицин высокоизбирательно ингибирует синтез рибосомной РНК и не влияет на синтез ДНК [Reich et al., 1962]. Использование радиоактивного дактиномицина показало, что он ковалентно связывается с остатком гуанина в ДНК и не соединяется больше ни с одним из компонентов клетки при концентрациях, блокирующих синтез РНК. Феноксазиновый цикл ин- теркалирует в ДНК около Г-Ц пары, а пептидная часть молекулы располагается в малой бороздке [Muller, Crothers, 1968] (подробнее об интеркаляции см. разд. 10.3.2).

Дактиномицин (актиномицин D) обладает ярко выраженным эффектом против опухоли Вильмса почки, одной из наиболее распространенных злокачественных опухолей у детей. Под его действием рассасываются даже легочные метастазы этой опухоли [Farber, Mitus, 1968]. При формах рака, требующих длительного лечения, дактиномицин не применяют в связи с его сравнительно невысокой избирательностью действия.
8-Азагуанин (4.40) впервые был получен синтетическим путем, однако позже оказалось, что ему идентичен антибиотик па- тоцидин, выделенный из Streptomyces albus [Anzai, Suzuki, 1961]. 8-Азагуанин применяют при опухолях мозга, почек и печени, так как в этих органах содержится много фермента гуана- зы, деаминирующей этот препарат до безвредного 8-азаксанти- на, который обычно отсутствует в клетках опухолей этих органов [Levine, Hall, Harris, 1963]. 8-Азагуанин, сильный ингибитор синтеза белка, не включается в ДНК Е. coli и бактериофага Т2 [Smith, Matthews, 1957], культуры опухолевых клеток или интактных мышей [Nelsen et al., 1975]. Гуанозин-5'-фосфатпиро- фосфорилаза превращает его в нуклеотид, включающийся в матричную РНК. Такая мРНК вызывает диссоциацию полисом до мономеров, останавливая тем самым синтез белка [Kwann, Webb, 1967]. 8-Азагуанозин-5-фосфат вмешивается в биосинтез пуринов, блокируя на ранних стадиях синтеза фермент фосфо- рибозилпирофосфатамидотрансферазу [McCollister et al., 1964].
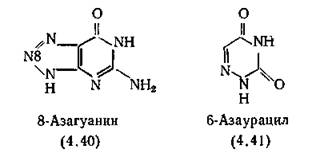
6-Азаурацил (4.41) используют в сельском хозяйстве в качестве фунгицида для борьбы с мучнистой росой (например, у огурцов). В клетках грибов он превращается в риботид, являющийся аналогом оротидиловой кислоты и блокирующий оро- тидилдекарбоксилазу [Dekker, 1968]. В медицинской практике он запрещен из-за побочного действия на ЦНС. Однако триаце- тилпроизводное этого риботида, азарибин (разд. 3.6), применяется при псориазе [Calabresi, Turner, 1966].
Митрамицин, производное тетрагидроантрацена с двумя присоединенными пиранозными кольцами, выделен из грибов Streptomyces. Он ингибирует синтез РНК, не влияя при этом на синтез ДНК- Митрамицин может вызвать гипокальциемию и в целом не очень избирателен, однако применяется при опухолях семенников [Hill et al., 1972].
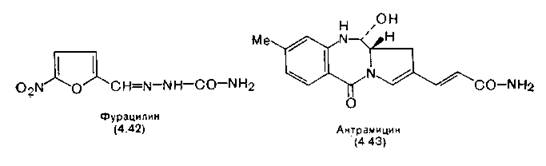
Фурацилин (4.42) (семикарбазон 5-нитрофурфурола) противобактериальное средство широкого спектра действия.
Эффек-тивен на последних стадиях трипаносомозов с резистентностью к препаратам мышьяка. В разбавленных растворах фурацилин ингибирует образование всех типов РНК в Е. coli [Tu, McCalla, 1976]. Он не действует на мутагенов, не имеющих «нитрофуран- редуктазы», что свидетельствует о том, что биологической активностью обладает восстановленная форма фурацилина (возможно, гидроксиламинопроизводное или его ближайший аналог). Фурацилин с потенциалом восстановления — 0,425 в (определен со стандартным каломельным электродом при pH 7) является акцептором электронов с высоким отрицательным потенциалом, находящимся на нижней границе биологически значимой области. Потенциал восстановления соответствующего производного бензола, семикарбазона пара-нитробензальдегида, равен —0,580 в, в связи с чем это соединение не восстанавливается в биологических системах [Sasaki, 1954]. В ряду нитрофуранов легче восстанавливаются соединения с сопряженной боковой цепью, так как образующийся, анион-радикал стабилизируется в этом случае сопряжением [Lindberg, 1970]. См. также разд. 5.4.3 (гидрогеносомы). К лекарственным веществам подобного типа относятся антисептик мочевых путей фурадонин (6.15) и метронидазол (6.22), производное нитромидазола, применяющееся при амебиазах и трихомонадозе.
Антрамицин (4.43), легко подвергающийся ковалентной гидратации (разд. 2.5), присоединяется к ДНК-связывающему месту РНК-полимеразы и ингибирует синтез РНК. Возможно, что именно с этим связано его противоопухолевое и антимикробное действие [Horwitz, Grollman, 1968; Kohn, Spears, 1970].
Гикантон (3.42,6), 4-гидроксиметил-1 - (2-диэтиламиноэтил- амино)-тиоксантен-9-он, применяют при шистосомозах. При однократном введении он мешает включению уридина в РНК клеток хозяина и червей, причем и чувствительных, и резистентных форм. Однако в клетках хозяина и резистентных форм червей этот эффект быстро пропадает, что и обеспечивает избирательность его действия [Mattoccia, belli, Cioli, 1981]. Гикантон обладает некоторыми побочными эффектами, например вызывает тошноту, а в модельных экспериментах показано его канцерогенное и мутагенное действие, в связи с чем не рекомендуется применять его в педиатрии или при риске повторного инфицирования.
Флаван (4.44), родоначальник бесчисленного множества окрашивающих веществ, содержащихся во фруктах и цветах, проявляет слабую активность против 20 штаммов обычных вирусов простуды в тесте бляшкообразования. Его высоколипофильный аналог — 4,6-дихлорпроизводное — более активен. Это соединение ингибирует синтез РНК в клетках паразита и обладает высоким терапевтическим индексом (мыши) [Bauer et al., 1981]. Однако эффективный способ введения такого водонерастворимого соединения в организм человека представляет собой проблему.
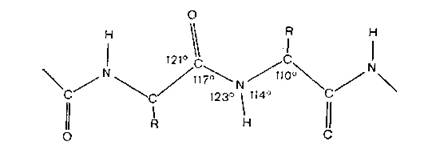
Рис. 4.2. Участок полипептидной цепи.
4.1. Белки
В любой живой клетке белки синтезируются в рибосомах приблизительно из 20 различных аминокислот L-конфигурации. ОММ белков варьирует от 6000 до миллиона и более, но все они имеют общую структуру (рис. 4.2), где R — довольно простые заместители, например метильная (для аланина) или пара-гидрок- сибензильная (для тирозина) группы. Регулярно издающийся в США Атлас последовательности и структуры белков (Atlas of Protein Sequence and Structure, Washington, D. С.) содержит сведения об аминокислотной последовательности для многих сотен белков (разд. 17.4).
Аминокислотная последовательность белка определяется генетической информацией, заложенной в клеточной ДНК, и реализуется через посредство мРНК и тРНК- Последовательность аминокислот в белке называется первичной структурой. Поли- пептидные цепи всегда образуют вторичную структуру, которая стабилизируется множеством водородных связей между группами —СО и —NH различных остатков. Изменение направления цепи происходит в месте расположения остатков пролина. Полипептиды образуют вытянутые цепи с расстоянием между аминокислотными остатками 0,72 нм. В природе из-за наличия боковых цепей у аминокислот неизбежно происходит некоторое искажение этой структуры, и поэтому периодичность составляет около 0,70 нм, как, например, в фиброине шелка.
При объемных боковых цепях в аминокислотных остатках вместо образования уложенных рядом цепей (^-структуры) происходит спирализация молекулы с образованием правозакрученной а-спирали, в которой на каждый виток приходится 3,6 ами-
нокислотных остатка. Как показали Payling и Corey, такая спиральная структура характерна для коллагена, в ней расстояние между аминокислотными остатками по оси составляет 0,15 нм,
а витки стабилизируются водородными связями СО----------- NH
между близрасположенными остатками аминокислот. Данные о вторичной структуре белка были получены методом рентгеновской кристаллографии с разрешением от 0,2 до 0,3 нм, определением дисперсии оптического вращения и числа атомов водорода, которые медленно обмениваются на дейтерий из D2O (т. е. участвующих в образовании межспиральных связей).
Эти спирали существуют в виде длинных нитей, которые в глобулярных (т. е. неволокнистых) белках (включая ферменты) складываются так, что получаются петли неправильной формы, характерные для третичной структуры. Форма петель обычно определяется аминокислотной последовательностью [Perutz, Kendrew, Watson, 1965]. Связи, стабилизирующие третичную структуру, могут быть ковалентными (дисульфидными), ионными, ван-дер-ваальсовыми и водородными. Например, активная конформация рибонуклеазы (белка, состоящего из 124 аминокислотных остатков) закреплена связями S—S между следующими парами аминокислотных остатков (считая от NH2 конца цепи): 28—84, 65—72, 40—95, 58—ПО. Молекула белка получается компактной, но содержит внутри несколько молекул воды. Почти все полярные группы (—ОН, ■—NH2, —СООН) обращены наружу, благодаря чему глобулярные белки растворимы в воде.
Рентгеноструктурное исследование кристаллов глобулярных белков показало, что структурная организация их молекул может быть трех типов: спирализованная, складчатая и беспорядочная. Беспорядочность структуры пептидной цепи может быть вызвана наличием остатков аспарагина, аспарагиновой кислоты и фенилаланина. Обычно кислые остатки расположены преимущественно около NH2 конца цепи, а основные (лизин, аргинин, гистидин) находятся ближе к СО2Н концу [Cook, 1967].
Некоторые пептиды синтезируются в живой клетке и без участия РНК, например трипептид глутатион в животных и растительных клетках. Бактерии без участия РНК могут синтезировать полипептиды, содержащие до 20 аминокислотных остатков.
Структура и функции ферментов рассматриваются в разд. 9.0. Более подробную информацию о структуре белков можно найти у Neurath, Hill (1975) и в периодическом издании Advances in Protein Chemistry. В приложении к книге «Структура и действие белков» [Dickerson, Geis, 1969] описан метод стереоскопической визуализации трехмерной структуры моделей белков.
4.1.1.