Дозирование жидкостей в асептических условиях — специфический процесс микробиологических производств
В настоящее время для промышленного микробиологического синтеза, осуществляемого в аппаратах периодического и непрерывного действия, характерно все бо- 168 лее широкое использование технических устройств и систем, предназначенных для дозированного введения различных стерильных жидкостей и чистых культур микроорганизмов.
Это обусловлено стремлением, с одной стороны, обеспечить большую стабильность поддержания параметров, определяющих состояние развивающихся культур микроорганизмов и их конечные характеристики, а с другой, исключить необходимость вмешательства операторов в процесс, доверив эту функцию автоматизированным системам регулирования и управления.Отмечается, что для большого класса микроорганизмов — продуцентов антибиотиков в практику все шире внедряются способы ферментации с непрерывным дозированием легкоокисляемых углеводов [26]. Однократная подача углеводов или даже подача раздельными порциями не обеспечивает достаточной скорости биосинтеза антибиотика (по сравнению с непрерывной подачей). Поскольку скорость подачи углеводов влияет на такой технологический параметр, как их концентрация в среде, естественно при изучении физиологии микроорганизмов находить зависимость между этим параметром и скоростью биосинтеза антибиотика. В реальных условиях, возникающих при непрерывном дозировании углеводов в ходе процесса, скорость их потребления настолько высока, что, несмотря на непрерывную подачу, в среде культивирования обнаруживают лишь следы углеводов. В то же время связь между скоростью дозирования углеводов и их концентрацией в среде неоднозначна и определяется концентрацией биомассы, ее возрастом, величиной pH и др. На примере пенициллина показано, что на продуктивность влияют микроконцентрации глюкозы в среде. Для роста характерна зависимость от концентрации глюкозы в виде кривой с насыщением. Продуктивность мицелия снижается при увеличении концентрации глюкозы.
Аналогичные данные применительно к процессу культивирования имеются не только по антибиотикам, но и по другим продуктам микробиологических производств. Во всех случаях использование дозированной подачи различных веществ обусловлено стремлением к повышению производительности стадии ферментации.Процесс, при котором в популяцию микроорганизмов непрерывно подаются питательные вещества без удале- 169
ния клеток, можно назвать циклически питаемым процессом культивирования. В определенных условиях получаемые культуры имеют характеристики, сходные с характеристиками культур, растущих в хемостате. Наиболее существенное сходство между двумя системами состоит в том, что удельная скорость роста клеток может регулироваться объемной скоростью подачи питательных веществ в ферментатор. В обеих системах, как правило, имеется один компонент питательной смеси, концентрация которого в среде определяет скорость роста. Другие компоненты могут присутствовать в количествах либо превышающих потребности клеток, либо субоптимальных, т. е. ниже такого значения, при котором один из компонентов становится фактором, почти ограничивающим рост. В хемостате в режиме установившегося состояния таким фактором является скорость разведения, которая регулирует концентрацию лимитирующего рост питательного вещества в среде, окружающей клетки. Таким образом, в хемостате численное значение удельной скорости роста определяется двумя величинами, не зависящими от общего количества клеток в сосуде для культивирования. Однако в некоторых ситуациях это положение справедливо и для циклически питаемых культур.
Предложены уравнения, устанавливающие связь между скоростью потока раствора питательных веществ и удельной скоростью роста микробов в циклически питаемой культуре [125]. Эти уравнения иллюстрируют основные свойства такого типа культивирования и демонстрируют возможные пути и условия проведения аналогии между ним и непрерывным культивированием. Различают три основных типа циклически питаемых культур: 1) культура, в которой удельная скорость роста микробов не меняется со временем; 2) культура, в которой скорость потока питательных веществ является функцией времени; 3) культура, в которой скорость потока питательных веществ постоянна.
В практических условиях возможны многочисленные вариации в режиме работы циклически питаемых культур. Например, непрерывная или периодическая подача питательных веществ: после определенного периода времени могут быть удалены либо клетки, либо использованная среда, а затем процесс выращивания можно по- 170
вторить или расширить. В работе [125] рассмотрены только непрерывно питаемые системы, при культивировании которых удаление клеток или жидкостей до окончания периода роста не производится. Выведенные уравнения соответствуют идеализированному состоянию микробного роста, во время которого выход клеток, рассчитанный по одному лимитирующему рост питательному веществу, остается постоянным. Поэтому предполагают, что все клетки находятся в состоянии сбалансированного роста, репликация клеток носит асинхронный характер, клеточный метаболизм не зависит непосредственно от возраста культуры и влиянием условий для поддержания роста можно пренебречь. Полагают, что увеличение объема среды в культуральном сосуде равно объему подаваемого питательного раствора.
Постоянная удельная скорость роста. Из уравнения экспоненциального роста микроорганизмов следует, что
Хд=Х„[ехр(рт) —11, (176)
где Хд—количество клеток в момент времени т=^0; Хо — то же в момент времени т=0; р— коэффициент скорости роста; т — время.
Кроме того,
= (177) где Vf — объем подаваемого в культуральный сосуд питательного раствора; — концентрация подаваемого лимитирующего рост питательного вещества; У^ — коэффициент выхода.
Комбинируя уравнения [176] и [177], получают;
V/=-Др--— [ехр (рт) — 1] (178)
у у
или
У/ = Ки[ехр(рт)-1], (179)
где значение Ку, которое называют «константой объема», представляет собой результирующий фактор, равный количеству клеток в момент времени т=0, деленному на произведение коэффициента выхода и концентрации лимитирующего рост питательного вещества в подаваемой среде.
Зависимость между скоростью потока питательного вещества и удельной скоростью роста получают путем дифференцирования по времени:
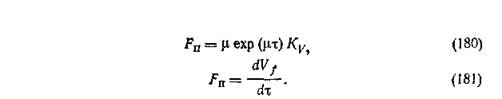
Постоянная скорость изменения потока.
Мгновенное значение скорости подачи питания выражается следующим уравнением:
где Fo — скорость потока лимитирующего рост питательного вещества при подаче в нулевое время; g— скорость изменения величины потока во времени.
Отсюда

Допуская, что подача питания является единственным фактором, влияющим на объем жидкости в культуральном сосуде, получаем:
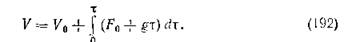
Следовательно,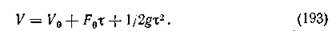
Постоянная скорость подачи питания. Когда скорость подачи питания поддерживается постоянной, значение g равно нулю и уравнение (191) приводится к следующему виду:
При т = 0 уравнения (191) и (194) становятся идентичными:
Аналогичным образом, приравнивая значение g к нулю, из уравнения (188) можно получить выражения для А и К.
Сравнение уравнений (176) и (178) показывает, что в случае циклически питаемого процесса с постоянной скоростью роста при определенных условиях концентрация клеток может поддерживаться постоянной. Это условие ограничивается тем, что константа объема Kv должна быть равна по величине объему жидкости в сосуде в нулевое время. То, что аналогичная ситуация имеет место при процессах, в которых скорость роста варьирует в зависимости от времени, подтверждается при сравнении уравнений (187) и (193).
Когда величина Kv равна Ко и скорость подачи питания постоянна, удельная скорость роста клеток в циклически питаемых культурах должна быть равна F/V—скорости разведения в сосуде для культивирования.
Это заслуживающее внимания условие получило название квази-установившегося состояния. Еще одно интересное состояние наблюдается, когда скорость экспоненциальной подачи определяется уравнением (180) и K.v равна Ко- В этом состоянии процесс культивирования с циклической подачей питания может осуществляться в течение какого-то периода времени с постоянными концентрацией клеток и удельной скоростью роста. Отме- 173тим, что предварительным условием для проведения любой достаточно близкой аналогии между циклически и постоянно питаемыми культурами должно быть равенство значений Kv и Vo- Однако процесс культивирования с циклической подачей питания необязательно проводить таким путем. Например, Kv может быть меньше значения Vo. Это возможно при подаче концентрированных питательных сред со сравнительно низкой скоростью в относительно большой объем содержимого в сосуде.
В табл. 37 приведены суммарные данные временной зависимости трех факторов, касающихся размножения
Таблица 37. Некоторые характеристики циклически питаемых процессов

клеток: удельной скорости роста р; скорости продуцирования клеток dXjdx и концентрации клеток X. Очевидно, что три основных типа циклически питаемого процесса: с повышающейся (экспоненциальной) скоростью подачи питательных веществ, с повышающейся (линейной) скоростью подачи питательных веществ и с постоянной скоростью подачи — обладают определенными свойствами. При процессе 1-го типа постоянна удельная скорость роста, а при процессе 3-го типа постоянна скорость продуцирования клеток. Исходя из равенства или неравенства Kv и Уо, можно судить, является ли концентрация клеток постоянной или нет при любом из трех типов процесса.
Для циклически питаемых процессов указанного выше типа характерны две особенности: 1) количество биомассы в ферментерах возрастает со временем; 2) постоянные удельная скорость микробного роста и проду-
цирование клеток являются взаимоисключающими.
В этих пределах существуют весьма разнообразные ус-, ловия, при которых может осуществляться выращивание микроорганизмов в циклически питаемых культурах. Возможность широкого использования этого метода культивирования иллюстрируется примерами, представленными в табл. 37. Несомненно, подобная универсальность является одним из основных факторов, определяющих использование циклически питаемых культур во многих производственных процессах.Показано, что скорость роста и концентрация подаваемого субстрата влияют на качество продукта, выходящего из хемостатов, которые содержат бактерии в чистой и смешанной культуре [115]. Таким образом, наметилось еще одно направление активного влияния на качество продукта с помощью непрерывного поддержания требуемой концентрации субстрата в культуральной жидкости. Решение этой задачи невозможно без использования дозирующих устройств.
При направленном биосинтезе одним из наиболее эффективных способов регулирования процесса является введение предшественников (процессы получения пенициллина и витамина В12) [6]. Доказана целесообразность введения предшественников в среду для ферментации новобиоцина. Исключительно важным является тот факт, что условия накопления антибиотика могут коренным образом отличаться от условий культивирования, при которых происходит накопление биомассы. Это предъявляет повышенные требования к процессам дозирования жидкостей, обеспечивающих регулирование процесса на обоих этапах.
9.2.