СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О МИКРОЦИРКУЛЯЦИИ, ПРОНИЦАЕМОСТИ КАПИЛЛЯРОВ И ТРАНСКАПИЛЛЯРНОМ ОБМЕНЕ
В процессе пренатального онтогенеза кровеносное русло дифференцируется на три отдела, выполняющих самостоятельные функции. Первый отдел — артериальный — обеспечивает доставку к органам и тканям пластических и энергетических субстратов.
Основной задачей второго отдела — венозного — является возврат крови к легким и сердцу и регуляция оттока, благодаря чему достигается тургор органов, реализуется депонирующая функция, осуществляется перераспределение крови. Наконец, третьим отделом являются сосуды местного (регио- иарного, органного) кровотока — артериолы, терминальные ар- териолы (метартериолы), капилляры, посткапиллярные венулы.Первые два отдела сосудистого русла, выполняющие, по сути дела, транспортную функцию, выделяют как центральное кровообращение, или макроциркуляцию. Третий отдел, непосредственно обеспечивающий питание тканей, определяют как периферическое кровообращение, или микроциркуляцию (Г. И. Мчед- лишвили, 1958; Zweifach, 1961). Ему принадлежит одна из важнейших функций системы кровообращения — доставка к клеткам паренхимы кислорода и питательных веществ, а также удаление из тканей продуктов обмена.
Такое деление кажется вполне целесообразным как с точки зрения биологической роли каждого из указанных отделов, так и особеппо их регуляции и саморегуляции.
По современным представлениям, система микроциркуляции включает сосуды диаметром от 2 до 200 мк (Maggio, 1965) и детально изучается в различных направлениях: морфологическом, биохимическом, физиологическом, патологическом, клиническом и др. Сам термин «микроциркуляция» возник недавно, в 1954 г., когда в США состоялась первая конференция по морфологии и физиологии микрососудов. Его рождение было связано с накоплением новых данных о строении, функции и циркуляторных расстройствах в системе кровеносных капилляров, которые было невозможно объяснить с позиций Krogh (1929).
Единого четкого определения микроциркуляции не имеется. В широком смысле под этим понимают закономерности цир-
куляции крови, лимфы и других биологических жидкостей в различных условиях жизнедеятельности организма (А. М. Чернух, 1968). Наиболее важным и изученным разделом ее является микрогемоциркуляция.
В представлении других (Fulton, 1962) микроциркуляция — это условия транспорта крови по микрососудам (артериолы — капилляры — венулы). Близкое определение дается в отечественной литературе (В. В. Парин, Г. И. Мчедлишвили, 1964); «Понятие „микроциркуляция" подразумевает не только собственно капиллярное кровообращение, но и функционирование других частей периферического русла, в той или иной степени оказывающих влияние на циркуляцию крови в капиллярах».
Микроциркуляторное ложе по своей организации и функции специфично для каждого органа, причем существует зависимость между строением капиллярного русла, током крови по капиллярам и функциями органа. Общей морфологической закономерностью микроциркуляторной системы является уменьшение количества мышечных элементов в стенках дистальных артериальных ветвей по мере приближения к капиллярной сети и нарастание мышечных элементов в стенках венозных ветвей по мере удаления их от капиллярной сети (В. В. Куприянов, 1969).
Микроциркуляторное ложе является той морфологической ОС- ПОВОЙ, где непосредственно совершается обеспечение органов и тканей всеми необходимыми питательными веществами и удаление из тканей продуктов их метаболизма. Однако сам переход веществ в направлении кровь — ткань и обратно есть явление более сложное и наряду с капиллярным кровотоком определяется рядом других морфологических структур, биологических и физико-химических закономерностей.
Таким образом, кровеносный капилляр, являясь частью замкнутой транспортной системы (сосудистой), в то же время приобретает принципиально новое качество. Последнее состоит в том, что капилляр входит как важная составная часть в систему трофического обеспечения тканей.
Возникновение и дальнейшее развитие капилляров в процессе фило-онтогенеза тесно связано с уложнением и специализацией клеточных элементов многоклеточного организма, требующих для своего существования и функционирования постоянных внешних условий, т. е. наличия в околоклеточной среде всех необходимых веществ. Это достигается, как известно, формированием так называемых тканей внутренней среды (мезенхимы), выполняющих трофическую роль (И. И. Мечников, 1892; А. А. Заварзин, 1945).
Дальнейшая дифференцировка и усовершенствование этой последней функции мезенхимы приводит к возникновению капилляров и подвижной трофической среды — циркулирующей крови с ее сложным аппаратом кроветворения.
Все сказанное позволяет представить капилляр как составную часть единой морфо-функциональной структуры, которая включает следующие образования: кровеносный капилляр с циркулирующей в нем кровью, перикапиллярные соединительнотканные элементы, лимфатические капилляры и прилегающие клетки паренхимы. Указанная морфофункциональная единица получила название капилляро-соединительнотканной структуры (А. И. Нестеров, 1929; В. П. Казначеев, 1960) или (для краткости) «микрорайона» (В. П. Казначеев, 1960; А. А. Дзизинский, 1970). Возникновение капилляро-соединительнотканных структур (микрорайона) в эволюции животного мира является одним из. важных приобретений в истории их развития. Основными функциями канилляро-соединительно- тканных структур являются защитно-барьерная и трофическая.
Защитно-барьерная функция. В начальный период изучения капилляров основное внимание было сосредоточено на выяснении гистоморфологических и физиологических особенностей. Вместе с тем подчеркивалась пограничная, барьерная роль кровеносных капилляров (Л. С. Штерн, 1935; Starling, 1909; Rrogh, 1929; Pappenheimer, 1963). Это нашло отражение и в терминологии, которой обозначались структуры, например «гисто-гематические барьеры» (Л. С. Штерн, 1935) или «гемато-паренхиматозные барьеры» (Н. Д. Стражеско, 1943; А. А. Богомолец, 1928).
В настоящее время защитнобарьерная функция капилляров изучена наиболее подробно (А. И. Нестеров, 1929; Л. С. Штерн, 1935; Г. Д. Залесский, 1949; Н. Д. Стражеско, 1955; В. П. Казначеев, 1960; А. Я. Росин, 1968; А. А. Дзизинский, 1970). Характерным ее проявлением служит различный уровень проницаемости капилляров в зависимости, с одной стороны, от их органной и тканевой принадлежности, с другой — от свойств проходящего вещества.Своеобразную защитную роль в ряде патологических ситуаций могут иметь и фибропластические (С. П. Шурин, В. П. Казначеев, 1964) или десмопластические (Ю. Г. Целлариус, 1964) процессы.
Трофическая функция капилляро-соединительнотканных структур состоит в обеспечении оптимальной среды для нормальной жизнедеятельности и функционирования органов и тканей. Указанные представления созвучны идеям А. А. Богомольца (1928) о трофической функции соединительной ткани и по сути дела могут рассматриваться как одно из конкретных выражений этой функции. Необходимо подчеркнуть, что под трофической функцией капилляро-соединительнотканных структур понимается не только транспорт и перемещение субстратов, но и соответствующая «переработка» их.
Эволюционно-сравнительные исследования, посвященные становлению системы трофического обеспечения многоклеточных организмов (И. И. Мечников, 1892; А. А. Заварзин, 1945), позволяют считать, что ферментативно-трофические превращения, начавшиеся в циркулирующей крови, являются начальными этапами процессов трофического обеспечения клеток паренхимы. С другой стороны, многочисленные ферментативные реакции, нередко возникая в перикапиллярных структурах, заканчиваются сложными биохимическими превращениями в циркулирующей крови.
Важно отметить, что в трудах И. И. Мечникова заложены также представления о биохимической природе трофической функции мезенхимы, первоначальную основу которой составляют процессы фагоцитоза и внутриклеточного пищеварения. В процессе эволюции многоклеточных организмов параллельно формированию капилляро-соединительнотканных структур возникали и те сложные ферментативно-биохимические механизмы и системы, которые составляют биохимическую основу их функции и обеспечивают трофический оптимум клеток паренхимы.
Это позволяет рассматривать «проницаемость» капилляро-соединительнотканных структур не только как проблему физиологии и патологии барьеров, но и как проблему функции сложной ферментативно-трофической системы, морфологической базой которой являются капилляро-перикапиллярные структуры и кровь.Таким образом, транскапиллярный обмен — это сложный биологический процесс, который определяется условиями микроциркуляции, фиксацией (сорбцией) субстратов, их перемещениями, превращениями и, наконец, захватом паренхиматозными клетками. Сказанное подтверждается данными изучения как пластического (белки, аминокислоты, полипептиды, нукле- протеиды и пр.), так и энергетического (гликоген, глюкоза, липопротеины, свободные жирные кислоты и пр.) обеспечения тканевых элементов.
Установлено, что азотистое питание клеток может происходить различно: во-первых, за счет аминокислот (так называемый аминокислотный тип питания), во-вторых, за счет пептидов и полипептидов («блочный» тип), причем используются пептиды, образованные из аминокислот как промежуточные этапы синтеза, и те пептиды, которые образуются при метаболизме белка. Полагают, что доля участия пептидов в построении белковых молекул должна быть значительной, ибо в термодинамическом отношении синтез белка из пептидов и полипептидов более выгоден, чем синтез белковых молекул непосредственно из свободных аминокислот (А. С. Коникова, М. Г. Крицман, 1965). В обоих случаях аминокислоты и полипептиды могут быть как плазмейного происхождения, так и образовываться в перикапиллярных пространствах при расщеплении белков крови, распадающихся клеток и межуточного вещества.
В связи с этим вопрос о проницаемости кровеносных капилляров для белков и других субстратов имеет принципиальное значение. Если сам факт выхода белков крови в ткани и обратно не вызывает сомнений, то трактовка физиологического значения этого процесса до последнего времени была неудовлетворительной. В настоящее время принято считать, что белки плазмы используются для построения клеточных белков (А.
С. Ко- никова, М. Г. Крицман, 1965; Whipple, 1956). Предполагали, что белки плазмы участвуют, даже без глубокого расщепления, в образовании специфических белков органов и тканей (Whipple, 1956). Однако этот взгляд не подтвердился. Наблюдения за распадом альбумина показали, что последний используется в качестве пластического материала лишь после предварительного распада до аминокислот или пептидных фрагментов (Walter е. а., 1957).Опираясь на собственные материалы клинических и экспериментальных исследований с учетом литературных данных, мы высказали предположение о том, что постоянное перемещение белков крови в перикапиллярные пространства является важнейшим звеном в трофической жизни паренхиматозных элементов (В. П. Казначеев, 1960, 1965).
Источником пластических субстратов, в том числе белка, могут быть также форменные элементы крови (лейкоциты), попавшие в ткани (Г. К. Хрущов, 1958). Внимание к лейкоцитам было привлечено классическими наблюдениями И. И. Мечникова, который определил их роль в воспалительном процессе. Впоследствии выяснилось, что лейкоцитарная реакция является универсальным, стандартным ответом на многие повреждения. Отсюда возникла мысль о том, что, помимо защитных функции, лейкоциты могут иметь определенное значение в восстановительном процессе. Эта мысль была развита Г. К. Хру- щовым, который обосновал концепцию стимуляционной функции лейкоцитов в восстановительном процессе. В обычных условиях лейкоциты стимулируют физиологическую регенерацию, компенсирующую рабочую изнашиваемость клеточных элементов. Основную роль в этих процессах, по данным Г. К. Хрущова, играют незернистые формы лейкоцитов (лимфоциты). На основании этих исследований были приготовлены лечебные лейкоцитарные сыворотки.
Вышедшие в перикапиллярное пространство белки крови, а также белки отмирающих клеток и форменных элементов крови подвергаются там ферментативному расщеплению до определенного уровня и усваиваются паренхиматозными клетками.
Многочисленные данные по распределению меченых белков и форменных элементов крови в организме человека и животных находятся в полном соответствии с указанным положением (Л. М. Тищенко и др.; 1967; Dykes, Jones, 1968). По данным ряда авторов, построение ядерного вещества растущими клетками в какой-то мере осуществляется за счет нуклеиновых соединений плазмы, а также распадающихся в тканях формен
иш элементов крови и других клеток (Policard, 1963). Из клеток крови источником нуклеиновых соединений в первую очередь являются лимфоциты.
«Процесс реутилизации отбросов при распаде лимфоцитов,— указывает А. Поликар,— представляет собой частный случай того биологического закона, что создание жизни интимнейшим образом связано с явлением смерти. Дегенерация лимфоцитов доставляет те неопределенные факторы, которые некогда были названы Carrel трефонами. Термин может быть и спорен, но самый факт неоспорим».
Не исключено, что наряду с трофической функцией нуклеиновые соединения лимфоцитов могут иметь информационное значение (Г. Д. Свет-Молдавский, 1968; Policard, 1963, и др.).
В последние годы появились сведения о том, что фибробласты, подобно клеткам нейроглии, могут служить источником выработки нуклеиновых соединений для паренхиматозных элементов.
Необходимо учесть также, что процессы доставки в клетки таких веществ, как медиаторы, гормоны, и других биологических веществ, осуществляющих регулирующую роль, также тесно связаны с функционированием капилляро-соединительнотканных структур.
Приведенные выше материалы убедительно свидетельствуют о том, что пластическое обеспечение клеточных элементов органов и тканей, основу которого составляют предшественники белков и нуклеиновых клеток, тесно связано с капилляро-соединительнотканными структурами как основным и неотъемлемым компонентом обменно-трофической сосудисто-тканевой системы.
Не меньшую роль, по-видимому, капилляро-соединительнотканные структуры играют и в доставке энергетических субстратов в ткани. Имеются данные о наличии на поверхности эндотелия капилляров липолитических ферментов и расщепления липопротеинов в пристеночном эндоплазматическом слое капилляров. По данным экспериментов на лягушках, поверхность эндотелиальных клеток капилляров обладает способностью расщеплять крахмал (А. М. Уголев, 1963). Фундаментальные исследования А. М. Уголева по эволюции пищеварения позволили предположить, что пристеночное пищеварение возникло раньше, чем появились специальные органы пищеварения. А указанные выше эксперименты дали основание автору поставить вопрос о том, не могут ли соматические клетки позвоночных животных, в частности эндотелий сосудов, в какой- то мере сохранить этот механизм. Не исключено, по мнению автора, что переход белков и полисахаридов, содержащихся в крови, через капилляры связан с их предварительным гидролизом. В этом случае мембранное пищеварение могло бы рассматриваться как один из элементов межуточного обмена.
И
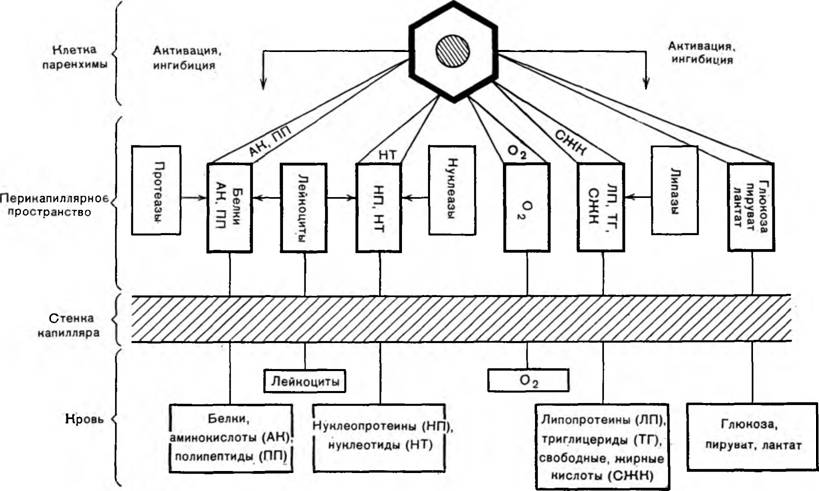
Трофическая функция капилляро-соединительнотканных структур не только включает в себя обеспечение клеточных элементов тканей, но, по-видимому, касается и межклеточного вещества. В образовании структурных элементов межклеточного вещества соединительной ткани могут использоваться некоторые углеводно-белковые комплексы сыворотки (М. С. Валитова, 1957). Для иллюстрации сказанного приведем схему трофического обеспечения клеток паренхимы (схема 1).
Обменно-ферментативные превращения, происходящие в капилляро-соединительнотканных структурах, наряду с трофическим обеспечением тканей имеют и другое фундаментальное значение — обеспечивают процессы репарации и восстановления тканей. Речь идет об участии системы свертывания крови и фибринолиза в процессах физиологической регенерации, а также восстановления ткани после повреждения (Astrup, 1968).
В обзоре Astrup (1968) приведены литературные и собственные данные автора, убедительно показывающие, что образование и растворение фибрина в перикапиллярных пространствах, которое происходит и в физиологических условиях, является важным фактором тканевого восстановительного процесса. В дополнение к уже известной роли фибрина как гемостатического барьера и ограничения экссудативного процесса, а также трофического белка фибрин формирует структуру для пролиферации и миграции клеток, а также представляет матрицу, на которой формируется репаратпвпая соединительная ткань. Однако по мере клеточной пролиферации фибрин должен лизироваться, так как в противном случае процессы репарации соединительной ткани закончатся формированием грубого рубца. Продукты расщепления фибрина, по-видимому, являются источником пластического обеспечения растущих клеток.
Таким образом, для оптимального восстановления ткани требуется динамическое адекватное соотношение между процессами образования и отложения фибрина и лизисом его. Нарушение этих соотношений будет приводить к различным расстройствам восстановительных процессов. Важно отметить, что фибрпполитическая активность в участках репарации обусловлена главным образом активаторами плазминогена, содержащимися в эндотелии капилляров.
Данные о распределении меченного фибриногена, а также плазмина (Amris е. а., 1964) в нормальных условиях и при различных патологических ситуациях согласуются с концепцией Astrup (1968).
Изложенные материалы убедительно показывают, что территория капилляро-соединительнотканных структур служит ареной чрезвычайно ответственных процессов по трофическому обеспечению клеток паренхимы и межуточной ткани. При этом капилляро-соединительнотканные структуры являются самыми активными участниками всех обменно-трофвгёеских процессов, начиная с циркулирующей крови и кончая ассимиляцией субстратов паренхиматозными элементами. Это дало нам (В. П. Казначеев, 1960) основание рассматривать «проницаемость капилляров» как активный процесс, в основе которого лежат ферментативные превращения, направленные на обеспечение оптимальной среды для жизнедеятельности паренхиматозных элементов. Этот процесс протекает в функционально единой структуре соединительной ткани (кровь, кровеносные капилляры, перикапиллярные клеточные и внеклеточные структуры, лимфатические капилляры, лимфа).
Таким образом, в нашем представлении такие понятия, как «трофическая функция капилляро-соединительнотканных структур» и «проницаемость кровеносных капилляров», являются тождественными, и в дальнейшем изложении мы будем руководствоваться сказанным. Однако в литературе имеются попытки рассматривать проницаемость капилляров более узко, как определенное морфологическое состояние структур самой капиллярной стенки — эндотелия, базальной мембраны и т. д. (И. А. Ойвин, 1966).
Во избежание терминологической путаницы можно пользоваться распространенным в иностранной литературе термином «транскапиллярный обмен», понимая под этим не простое перемещение (транспорт) субстратов, а сложный биологический процесс, идентичный приведенному выше понятию «проницаемость капилляров». Именно в таком понимании мы и будем пользоваться термином «транскапиллярный обмен».
Представление о проницаемости (транскапиллярном обмене) как активном физиологическом процессе, обеспечивающем оптимальное постоянство состава и свойств непосредственной внутренней среды клеток, тканей и органов, отражает по существу главную биологическую, физиологическую особенность изучаемого процесса, не ограничивая средства и механизмы, с помощью которых эта главная цель достигается.
Три основных механизма транспорта веществ — диффузия, фильтрация и активный перенос — обусловливают выполнение основной физиологической роли капилляро-соединительнотканных структур — обеспечение оптимального постоянства непосредственной внутренней среды клеток и тканей. Однако современный уровень методических возможностей, особенно при наблюдении за человеком, не позволяет четко расчлепить и выделить удельный вес таких факторов, как гидростатическое н онкотическое давление, скорость капиллярного кровотока и лимфотока, число функционирующих капилляров, изменение самих капилляро-соединительноткапных структур и зависящих от них величин диффузии, фильтрации и активного переноса. Чаще всего мы располагаем валовыми показателями перехода жидкости и субстратов из крови в ткани и обратно, по которым її судим о процессе проницаемости в целом, вычисляя так называемую суммарную проницаемость (И. А. Ойвин, 1966).
Потребность клеток в пластических и энергетических субстратах в каждый данный момент определяется их функциональным состоянием, поэтому должны существовать тонкие механизмы регуляции указанных процессов.
Факторами, определяющими транскапиллярный обмен, являются в первую очередь гемодинамика капиллярного кровотока (скорость кровотока, количество функционирующих капилляров, степень их дилятации, гидростатическое давление и т. д.), состояние самих капилляро-соединительнотканных структур, т. е. их проницаемость в узком понимании и, наконец, характер вещества, проходящего из крови в ткани и обратно. Изменения проницаемости капилляров (транскапиллярного обмена), а следовательно, и ее регуляция, возможны при направленном воздействии на каждый из указанных компонентов.
На современном уровне знаний система регуляции процессов проницаемости капилляров представляется довольно сложным механизмом, включающим нервные, гормональные и гуморальные звенья. В своем единстве и взаимосвязи указанные звенья составляют тот механизм регуляции, который обозначается как нейро-гуморальный.
Нейро-эндокринное звено механизмов регуляции процессов проницаемости капилляров поддерживает общий «настрой» их, являясь как бы «стратегическим управлением», в то время как непосредственное приспособление уровня проницаемости к трофическим нуждам каждого органа и ткани обеспечивается местными гуморальными веществами, так называемыми тканевыми гормонами, и рядом ферментов крови и тканей («тактическое управление»).Микрососудистая сеть, включая истинные капилляры, имеет непосредственный контакт с окружающими их клетками паренхимы и соединительной ткани и, несомненно, испытывает влияние различных химических веществ, выделяемых этими клетками. По представлениям Zweifach (1961), на этом уровне сосудистого русла создается третий регуляторный механизм — химические вещества, выделяющиеся в межуточные пространства тканей в процессе клеточного обмена. Среди последних наибольшее значение имеют вазоактивные амины и полипептиды — гистамин, серотонин, адреналин, ацетилхолин, кинины, некоторые ферменты крови и тканей — муколитические, протеолитические и др., биологически активные вещества — гепарин и др.