Метод внеклеточной регистрации электрических явлений при возбуждении одиночного мышечного волокна
Регистрация проводится с помощью двух электродов, расположенных на поверхности клетки и соединенных с гальванометром, регистрирующем разность потенциалов между двумя электродами.
Один электрод соединяют при этом с положительным полюсом гальванометра (активный или дифферентный электрод), а другой - с отрицательным (референтный электрод). Гальванометр соединен с пишущим устройством, осуществляющим графическую запись электрических явлений.В состоянии покоя одиночного мышечного волокна, наружная поверхность которого положительно заряжена, электрические явления не регистрируются (состояние статической поляризации) (рис. 3). Графически этот состояние представлено прямой линией - изолинией.
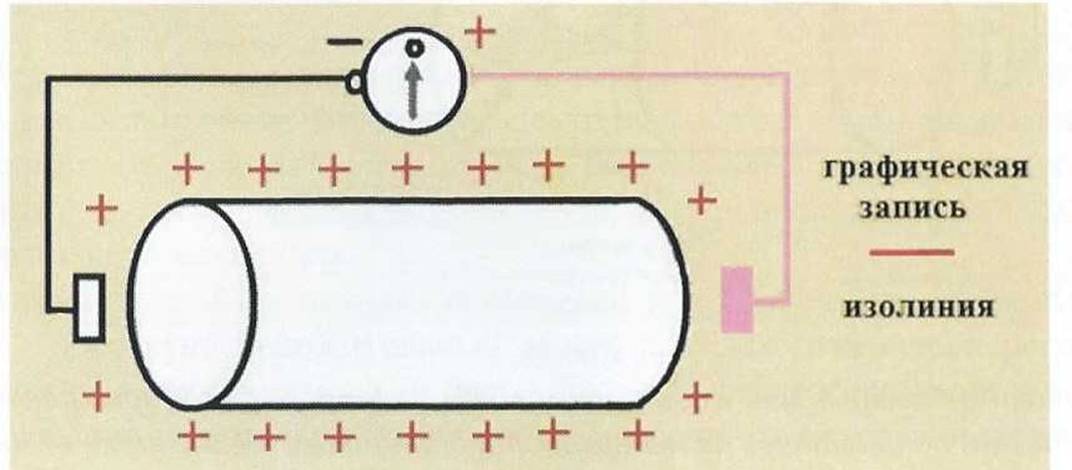
Рис . 3.
Состояние статической поляризации в одиночном мышечном волокне в состоянии покоя.
При возбуждении клетки (рис. 4) из точки воздействия возбуждающего электрического, химического или механического импульса начинается перезарядка (деполяризация) наружной поверхности цитоплазматической мембраны, в связи с изменением ее проницаемости к ионам.
Возбужденные участки мембраны, находящиеся в состоянии деполяризации, заряжены отрицательно, а не возбужденные - положительно. Появляется разность потенциалов и образуется система, состоящая из двух равных по величине, но противоположных по знаку зарядов, называемая диполем (рис. 5). Между отрицательным и положительным зарядами находится нулевая линия, на которой величина заряда равна нулю. Рядом с нулевой линией со стороны отрицательного поля расположен наибольший отрицательный заряд и наибольший положительный заряд. Вокруг положительного заряда располагаются силовые линии положительного поля, вокруг отрицательного заряда расположены силовые линии отрицательного поля. Появляется электродвижущая сила (ЭДС), величина которой регист-
рируется гальванометром и изображается графически.
Появляется ЭДС, которая является векторной величиной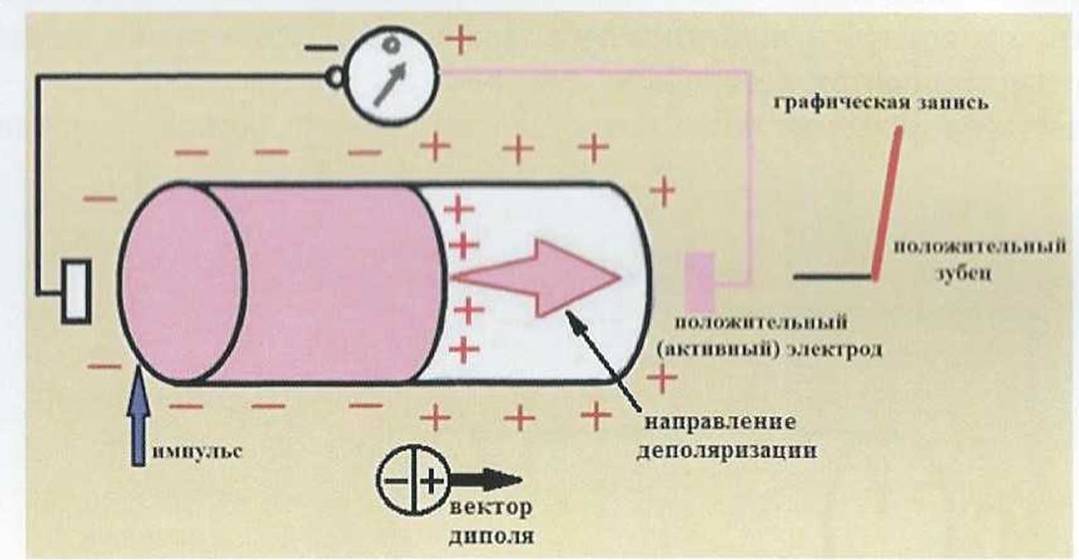
Рис. 4.
Начало деполяризации в одиночном мышечном волокне. Формирование разности потенциалов на поверхности одиночного мышечного волокна при его возбуждении и графическая регистрация процесса деполяризации. Здесь и на рисунках 6,7 красным цветом показаны возбужденные участки клетки. Здесь и на рис. 46 стрелкой указан участок клетки, на который оказывается возбуждающее воздействие.
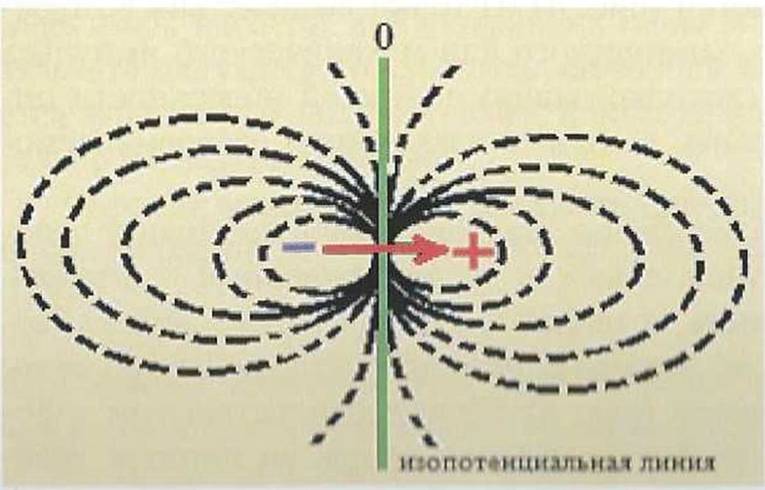
Рис. 5. Силовые линии электрического поля. Стрелкой показано направление вектора ЭДС.
)Д( является пік горі юн величиной, т.е. она имеет не только абсолютне и і.і• и-m її но н направление. Вектор ЭДС изображается в миле Oiречка прямой определенного масштаба со стрелкой. Условно прппяіо CMirnriь. пт стрелка направлена от отрицательного к поло-
9
жительному значению диполя, т.е. от возбужденного к невозбужденному участку клетки.
Конструктивно пишущее устройство в случае направления стрелки вектора ЭДС (на электрод направлен положительный заряд диполя) на активный электрод графически изображает отклонение вверх от изолинии - положительный зубец, а в случае направления стрелки от активного электрода (на электрод направлен отрицательный заряд диполя) отклонение вниз от изолинии - отрицательный зубец. Величина регистрируемого гальванометром тока и, соответственно, зубцов зависит от расстояния, на котором располагается изопотенциал ь- ная линия диполя от активного электрода (обратно пропорциональна квадрату расстояния). Чем дальше расположена изопотенциальная линия диполя от активного электрода, тем меньше влияние ЭДС и меньше величина зубца.
На рисунке 4 на активный электрод направлена стрелка вектора ЭДС, действует положительный заряд, поэтому графически регистрируется положительный зубец. По мере распространения волны возбуждения (деполяризация) вдоль мышечного волокна и приближения изопотенциальной линии диполя к активному электроду амплитуда зубца будет возрастать.
Когда все волокно окажется в состоянии возбуждения (рис. 6) и вся его поверхность будет заряжена отрицательно (статическая поляризация), разность потенциалов между электродами снова окажется равной нулю и будет записываться изолиния.
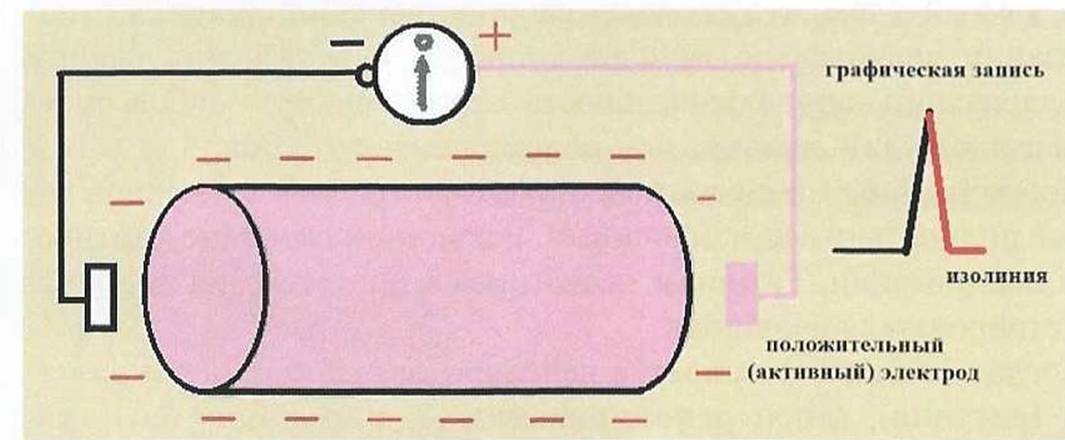
Рис. 6.
Одиночное мышечное волокно в состоянии возбуждения (состояние
статической поляризации).
Процесс реполяризации одиночного мышечного волокна начинается из той же точки, что и волна деполяризации (рис. 7). При этом часть мышечного волокна, вышедшая из состояния возбуждения, заряжается положительно, а часть волокна все еще находящаяся в состоянии возбуждения, заряжена отрицательно. Вновь возникает диполь и ЭДС, вектор которой направлен от активного электрода. К активному электроду обращена поверхность с отрицательным зарядом.
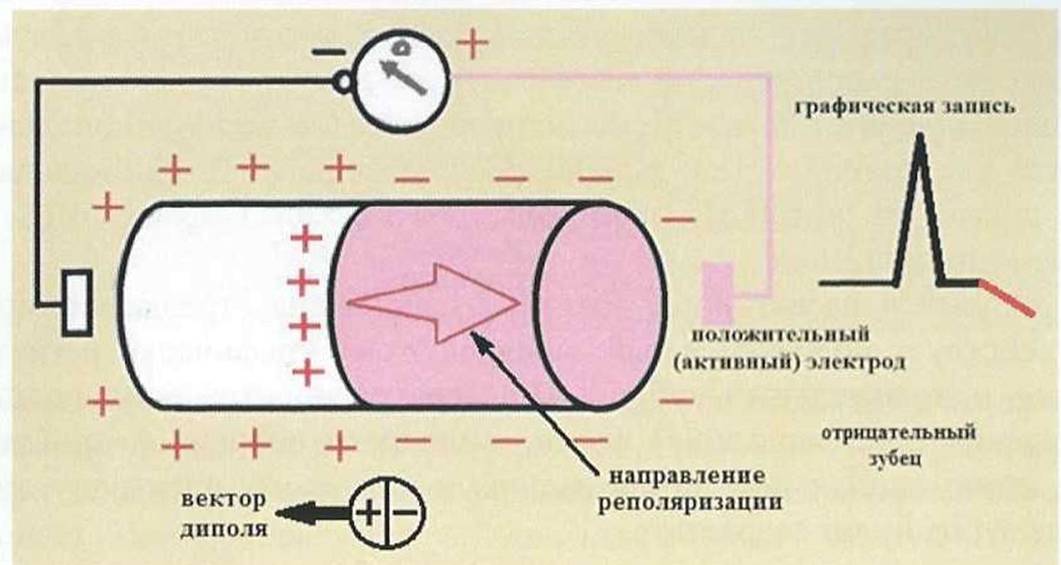
Рис. 7.
Реполяризсщия в одиночном мышечном волокне.
Графически регистрируется отрицательный зубец. Кроме того, в связи с тем, что скорость распространения процесса реполяризации значительно меньше скорости перемещения фронта деполяризации, продолжительность отрицательного зубца больше, а амплитуда - меньше, чем положительного зубца.
По окончанию процесса реполяризации (рис. 8) мышечное волокно полностью восстанавливает исходное состояние статической поляризации. Разности потенциалов при этом нет и будет регистрироваться изолиния.
Когда вся мышечная клетка деполяризована, клетка сокращается (систола), а при реполяризации — расслабляется (диастола).
На форму регистрируемых зубцов при внеклеточном отведении влияет не только электрическая активность самого мышечного волокна, но и место расположения активного электрода (смотри При-
ложение). При внутриклеточном отведении форма регистрируемо потенциала не зависит от места прокола мембраны.
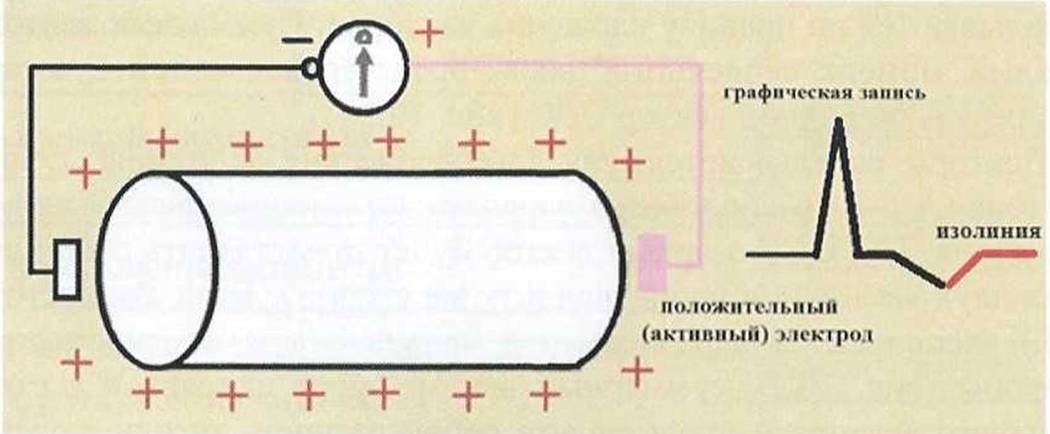
Рис.
8.Возврат мышечного волокна в состояние покоя (статическая
поляризация).
На рисунке 9 представлено сопоставление графических изображений процессов деполяризации (возбуждения) и реполяризации одиночного мышечного волокна при внутриклеточной и внеклеточной регистрации с указанным выше расположением электродов.

Рис. 9.
Графическая запись процесса возбуждения одиночного мышечного волокна.
А - внутриклеточное отведение; Б - внеклеточное отведение.
В сердце каждое возбужденное волокно представляет собой диполь, вызывающий появление ЭДС. В сердце одномоментно существует огромное количество диполей. Векторы возбуждения этих диполей складываются по правилу сложения векторов. Сумма всех векторов в каждый момент сердечного цикла представляет собой суммарный или результирующий вектор ЭДС (рис. 10).
Векторы складываются, как алгебраические величины. Если два вектора (А и В) расположены параллельно и направлены в одну сторону (рис. 10.1). Суммарный вектор будет представлять собой сумму этих двух векторов и направлен в ту же сторону. Если два вектора (А її Б) расположены параллельно и направлены в противоположные стороны (рис. 10.2), суммарный вектор будет направлен в сторону большего вектора и представлять собой разность между двумя векторами: из большего вектора (А) вычитается меньший вектор (Б). Если два вектора (А и В) направлены под углом друг к другу (рис. 10.3), их располагают так, чтобы они исходили из одной точки. Затем, считая, что такие векторы образуют две стороны параллелограмма, достраивают параллелограмм и проводят диагональ из точки, где совмещены начала обоих векторов. Диагональ параллелограмма представляет собой сумму двух векторов. В случае если два вектора равны по величине и направлены в противоположные стороны (рис. 10.4), то суммарный вектор будет равен нулю.
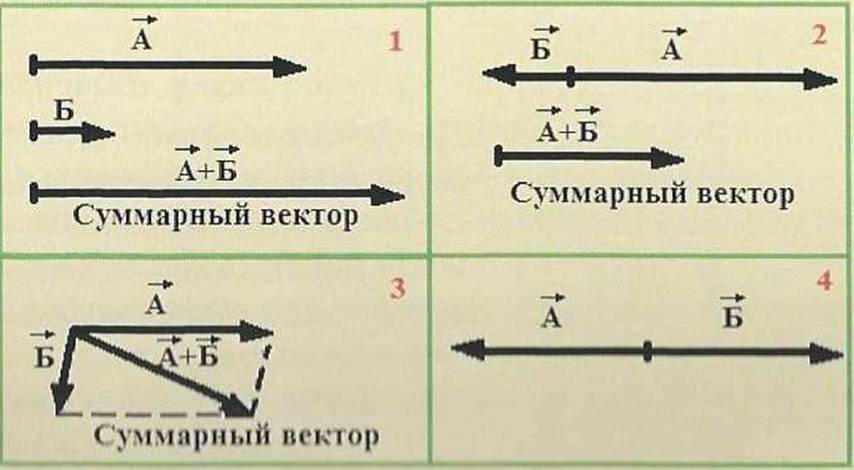
Рис, 10.
C 'ложение нектаров.
/ два вектора паправлепві в одну стороны; 2— два вектора направлены в противоположные стороны; 3— два вектора направ к ■///./ под г. 'лом друг к другу; 4 два вектора равны по величине н направлены в противоположные стороны.
Мышца сердца состоит из клеток двух видов - клеток проводящей системы и сократительного миокарда.
Сердце обладает четырьмя основными функциями, определяющих особенности его работы:
Функция автоматизма — это способность сердца вырабатывать электрические импульсы при отсутствии внешних раздражений.
Функцией автоматизма обладают только клетки синусового узла (пейсмекеры (от англ, pacemaker - водитель), P - клетки)и проводящей системы предсердий и желудочков. Сократительный миокард лишен функции автоматизма.
В основе автоматической активности Р-клеток лежит их способность к спонтанному медленному уменьшению величины МП — уменьшению отрицательного заряда внутренней поверхности мембраны во время диастолы (рис. 11). Этот процесс получил название спонтанной медленной диастолической деполяризации. При достижении критического уровня деполяризации развивается быстрая деполяризация с перезарядкой мембраны и появлению электрического импульса. Синхронное возбуждение Р-клеток ведет к суммации электрических импульсов и распространению этого импульса по проводящей системе миокарда.

Рис. 11.
Процесе самовозбуждения P-клетки. Внутриклеточное отведение.
Различают три центра автоматизма (рис. 12):
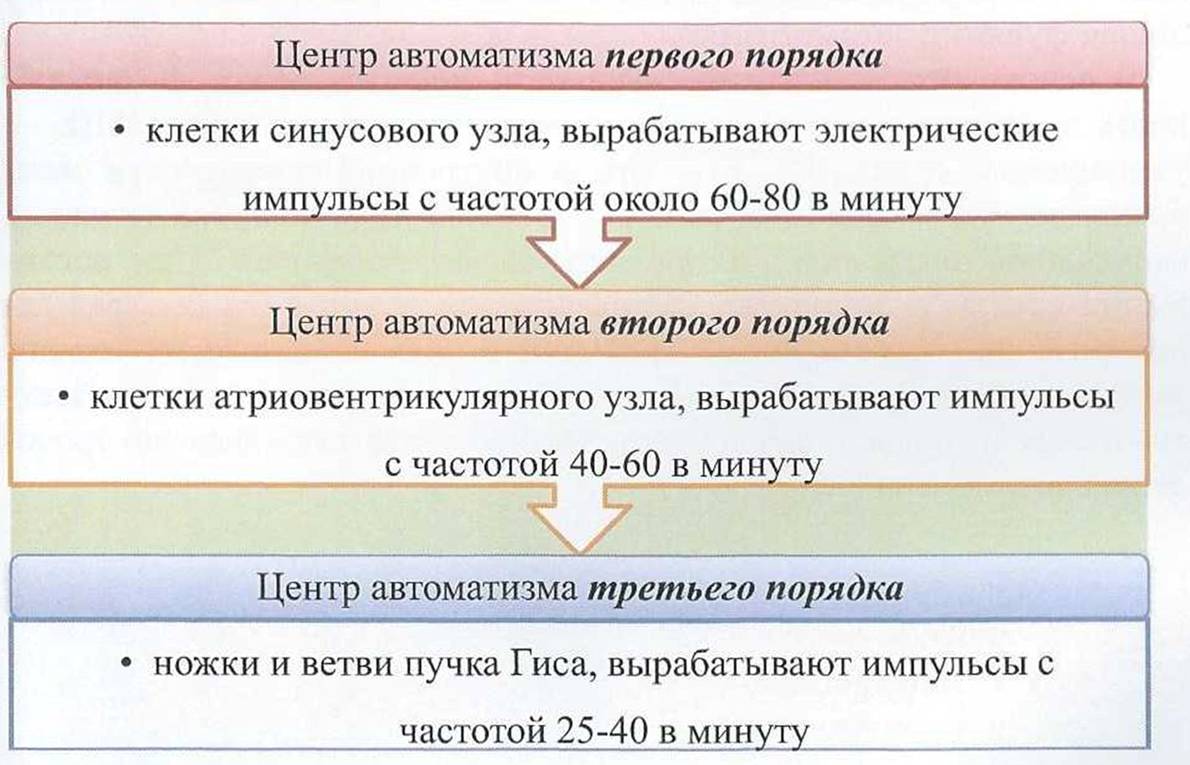
В норме единственным водителем ритма является синусовый узел, который подавляет автоматическую активность остальных (эктопических) водителей ритма.
При поражении синусового узла или нарушении проведения возбуждения к атриовентрикулярному узлу, последний становится водителем ритма.
На функцию синусного узла и других водителей ритма большое влияние оказывает симпатическая и парасимпатическая нервная система: активация симпатической системы ведет к увеличению автоматизма клеток синусового узла и проводящей системы, а парасимпатической системы - к уменьшению их автоматизма.
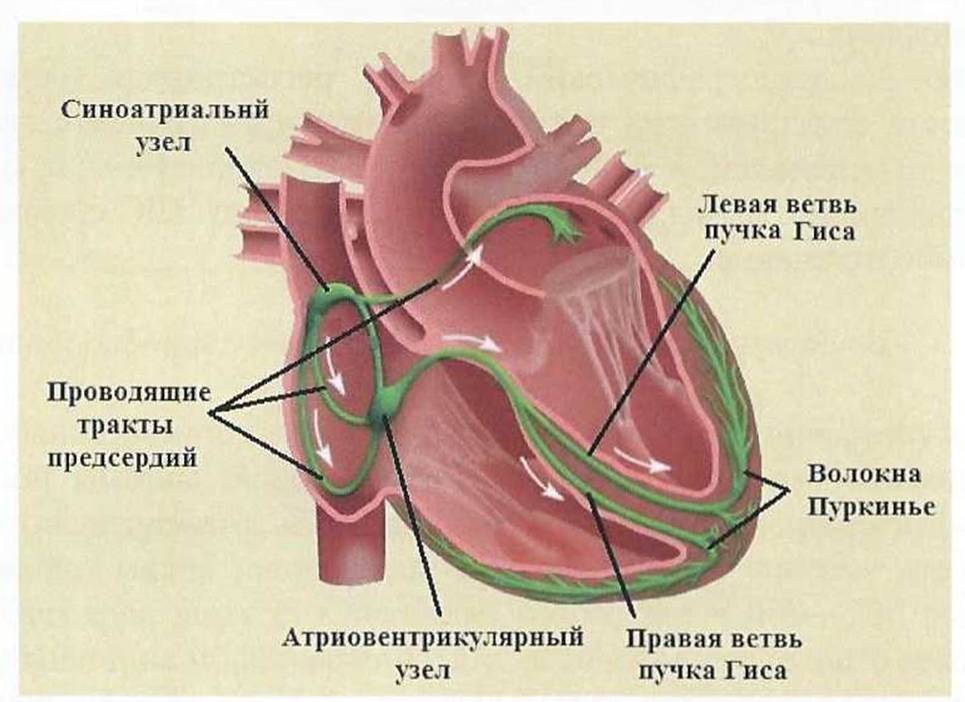
Рис. 12.
Проводящая системы сердца.
Возбудимость - это способность сердца возбуждаться под влиянием импульсов.
Функцией возбудимости обладают клетки как проводящей системы сердца, так и сократительного миокарда. Возбуждение сердечной мышцы сопровождается, возникновением ПД и в конечном счете - электрического тока.
Проводимость -это способность к проведению возбуждения, возникшего в каком-либо участке сердца, к другим отделам сердечной мышцы.
Сократимость - это способность сердечной мышцы сокращаться в ответ на возбуждение. Jrnoii функцией в основном обладает сократительный миокард (миокардиоциты) и нс обладают клетки водителей ритма проводящей системы.
В результате последовательного сокращения различных отделов сердца и осуществляется основная - насосная - функция сердца.
Суммарный или результирующий вектор ЭДС сердца, формирующийся при возбуждении клеток сердца можно зарегистрировать с поверхности тела при помощи электрокардиографического метода исследования.
Электрокардиографический аппарат регистрирует напряжение (разность электрических потенциалов) между 2 точками, то есть в каком-то отведении, и фиксирует на бумаге (экране) в виде кривой электрокардиограммы (ЭКГ) величину проекции ЭДС сердца на какое-либо отведение.
2.