ВВЕДЕНИЕ
Интуитивно ясно, что биогеоценоз, экосистема, биологическое сообщество, существующее в более или менее неизменном виде достаточно длительное время, обладают некоторой внутренней способностью противостоять возмущающим факторам, которые в изобилии поставляет внешняя среда (в том числе и человек).
Эту способность экологической системы обычно называют «устойчивостью» или «стабильностью». Несмотря на кажущуюся очевидность понятия, дать ему четкое и однозначное определение оказывается трудной задачей. До сих пор этот сильно перегруженный термин не имеет установившегося («устойчивого») определения [1]).Анализируя самые разнообразные высказывания по этому вопросу [2]), можно выделить некоторые группы требований к различным биологическим системам — требований, объединенных одним словом «устойчивость».
Первое — это требование известной неизменяемости во времени целого географического региона или ландшафта. Регион может включать в себя большое число различных биогеоценозов, а общая экосистема региона — состоять из разных экосистем, достаточно слабо связанных друг с другом. Основными процессами, определяющими динамику региона, будут не изменения численности населяющих его отдельных видов, а, скорее, глобальные биогеохимические циклы.
Вторая группа требований — сохранение числа видов в данном биологическом сообществе. Биологическое сообщество можно рассматривать как более высокий, чем популяционный, уровень организации живого вещества и определить как некоторую совокупность популяций, населяющих определенную территорию и образующих определенную структуру пищевых (трофических) связей и метаболизма. Концепция биологического сообщества отражает тот факт, что популяции живых организмов не разбросаны случайным образом по Земле как независимые группы, а образуют организованные системы. Сообщество считается устойчивым, если число составляющих его видов не меняется в течение достаточно длительного времени.
Пожалуй, именно это экологическое определение ближе всего к различным математическим определениям устойчивости, и именно по этой причине в данной книге рассматриваются проблемы устойчивости в основном биологических сообществ.И, наконец, третья группа требований относится скорее к отдельным популяциям, чем к сообществам. Считается, что сообщество устойчиво или стабильно, если численности составляющих его популяций не испытывают резких колебаний. Это определение более близко к физическому (точнее, термодинамическому) понятию стабильности системы. В термодинамике система считается стабильной, если малы вероятности больших флюктуаций, которые могут увести ее далеко от равновесного состояния и даже разрушить. Очевидно, что общие термодинамические концепции (например, связанный со вторым законом термодинамики принцип стабильности) должны быть приложимы и к биологическим (в частности, к экологическим) системам. Согласно этим концепциям любая замкнутая система с протекающим через нее потоком энергии (будь то биосфера или маленькое озеро) с большей вероятностью развивается в сторону некоторого устойчивого состояния, и в ней с необходимостью должны вырабатываться саморегулирующие механизмы. Когда это состояние достигнуто, то перенос энергии обычно идет в одном направлении и с постоянной скоростью, что соответствует термодинамическому принципу стабильности. Классическим примером такой проточной системы является система хищник — жертва. К сожалению (и в этом свойство всех слишком общих концепций), термодинамический принцип стабильности не дает нам метода, который позволил бы эффективно судить о степени стабильности конкретных биологических сообществ или экосистем.
С другой стороны, существует развитая математическая теория устойчивости (со всеми ее многочисленными приложениями в науке и технике), в которой определение устойчивости дается совершенно строго. Но все дело в том, что эта теория работает не с самими реальными объектами, а с их математическими моделями.
Поэтому, если мы имеем достаточно «хорошую» (в смысле адекватности и полноты описания) математическую модель биологического сообщества или экосистемы (например, в терминах дифференциальных или разностных уравнений), то на вопрос об устойчивости реального сообщества можно ответить, исследуя нашу модель обычными методами теории устойчивости. "Однако и в математической теории устойчивости существуют различные определения этого понятия — это уже другой вопрос. Обычно говорят об устойчивости некоторых решений системы уравнений, т. е. об устойчивости того или иного состояния системы или об устойчивости (стабильности) того или иного режима ее функционирования. Например, можно считать, что сообщество или экосистема устойчива, если траектория ее модели в фазовом пространстве не будет выходить за пределы заданной ограниченной области при некоторых возмущениях достаточно широкого спектра. Подобное определение отвечает интуитивным представлениям экологов и иногда является достаточным для них. Правда, мы здесь совершаем подмену, заменяя сам объект его математической моделью и оставляя в стороне вопрос о мере адекватности модели моделируемому объекту. Гносеологически такой подход вполне оправдан, ибо анализ устойчивости модели позволяет формулировать различные гипотезы о закономерностях функционирования моделируемого объекта, наличие либо отсутствие которых в реальности дает основание судить в том числе и об адекватности модели.Принципиальной альтернативой данному подходу является подход, основанный на попытках связать устойчивость сообщества с какой-либо другой реально (и достаточно просто) измеряемой его характеристикой. Среди экологов считается почти аксиомой, что более сложные по своей структуре, более богатые по числу ВХОДЯЩИХ в них видов сообщества и более устойчивы. По-видимому, это объясняется следующим. Различные виды по-разному приспособлены к изменениям окружающей среды. Поэтому широкий набор видов может реагировать на разнообразные изменения внешней среды успешнее, чем сообщество, состоящее из малого числа видов, а следовательно, первое устойчивее, стабильнее второго.
Вероятно, этим и обусловлено использование различных мер видового разнообразия (в частности, информационной энтропии или некоторых ее аналогов) в качестве характеристики устойчивости сообщества. Наиболее популярна информационная мера разнообразия (по Шеннону)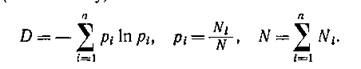
Здесь п — число видов в сообществе, Nt — численность і-го вида. Или несколько другая мера:
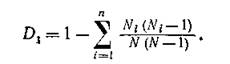
Можно показать, что D± — это вероятность того, что при случайной выборке по два из совокупности N индивидуумов они не будут принадлежать одному и тому же виду.При больших N и достаточно равномерном распределении по видам 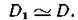 В самом деле,
В самом деле,
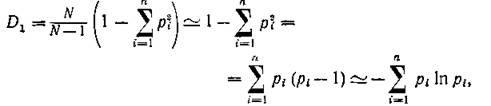
так как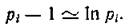 Предлагались и другие определе
Предлагались и другие определе
ния меры устойчивости (например, использующие предложенную Р. Фишером меру информации), но все они, по сути дела, используют понятия, первоначально возникшие в физике или теории информации.
Логично предположить, что в состоянии равновесия сообщество максимально стабильно и, следовательно, должно обладать максимальным разнообразием. Но, как легко показать, этому соответствует такая структура сообщества, при которой особи любых видов встречаются с одинаковой частотой (max D достигается при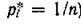 і, —
і, —
все виды одинаково обильны, нет доминирующих видов, в сообществе не существует количественной иерархии. Но наблюдения над реальными сообществами (в частности, над наиболее естественно выделяемыми элементарными составляющими биосферы — биогеоценозами) говорят совсем о другом — большинство достаточно долго существующих (а значит, и устойчивых) сообществ содержит доминирующие виды, которые осуществляют основную работу по переработке вещества и энергии в сообществе, т.
е. сообщества имеют иерархическую структуру. Это наводит на мысль, что использование в качестве меры устойчивости сообщества его разнообразия не совсем оправдано.Заметим однако, что во многих лабораторных (и не только лабораторных) сообществах на ранних стадиях их эволюции при движении к равновесному состоянию наблюдается увеличение разнообразия (увеличение D). По- видимому, эта мера все же в какой-то степени характеризует сообщество; более того, она, по всей вероятности, может даже характеризовать его устойчивость, но не всюду, а лишь на ранних стадиях эволюции.
В чем же причина этих парадоксов? Скорее всего, в формальном применении моделей физики и теории информации к системам, к которым они неприменимы. Как больцманов- ская энтропия в статистической физике, так и информационная энтропия в теории информации имеют смысл лишь для ансамблей из слабо взаимодействующих частиц или каких- либо других объектов. Введение энтропийной меры для таких множеств вполне обосновано. Но как только мы имеем дело с системами, элементы которых сильно взаимодействуют между собой, энтропийная мера неудовлетворительна. А биологические сообщества, где конкурентные взаимоотношения наиболее сильно проявляются вблизи положения равновесия и вся структура которых в основном определяется не характеристиками, присущими собственно виду, а характеристиками межвидовых взаимоотношений, представляют собой именно системы с сильными взаимодействиями.
С этой точки зрения понятны удачи в применении энтропийных мер на ранних стадиях эволюции сообществ. Все дело в том, что на этих стадиях, вдали от положения равновесия, конкуренция еще слаба, конкурентные давления малы, и сообщество вполне может рассматриваться как система со слабыми взаимодействиями.
Как видно из всего сказанного выше, на вопрос о причинно-следственной связи между разнообразием сообщества и его устойчивостью нельзя ответить однозначно. Хорошо говорит об этом Ю. Одум [3]): «...Следует решить, представляет собой разнообразие только «приправу» к жизни или оно необходимо для долгой жизни всей экосистемы, в которую входят и человек, и природа».
До сих пор мы никак не определяли, что такое устойчивость сообщества, ограничиваясь лишь интуитивным пониманием (по этой причине мы использовали понятия «устойчивость» и «стабильность» как эквивалентные). Но если мы хотим двигаться дальше, то нам необходимо дать этому понятию формальное (и в то же время конструктивное) определение.
Мы будем говорить, что сообщество устойчиво, если устойчиво некоторое нетривиальное положительное решение системы дифференциальных (разностных, дифференциально-разностных и т. д.) уравнений, являющейся моделью этого сообщества. В дальнейшем нас будет интересовать, в основном, устойчивость только стационарных решений, причем таких, для которых численность ни одного из видов не равна нулю или бесконечности. Эти решения мы будем называть нетривиальным равновесием (а зачастую и просто равновесием) сообщества. Из устойчивости таких решений следует сохранение числа видов в сообществе (обратное, вообще говоря, не обязательно). Таким образом, мы формализовали одно из экологических определений устойчивости.
Вполне естественно, что исключение одного или нескольких видов из сообщества можно интерпретировать как нарушение его устойчивости. Оставшиеся виды могут образовать устойчивое сообщество, но оно уже будет другим.
Ранее мы уже говорили о том, что в математической теории устойчивости существует много определений ЭТОГО понятия. Одно из основных — это устойчивость по Ляпунову. Именно это понятие будет наиболее часто использоваться в данной книге. Поэтому мы предполагаем, что читатель в той или иной степени уже знаком с основными идеями и методами ляпуновской теории устойчивости.
Однако здесь будут использоваться идругие определения устойчивости. Например, устойчивость, понимаемая как ограниченность решения сверху и снизу (ее часто называют устойчивостью по Лагранжу}. Для чего это нужно, поясним на примере. Пусть численности видов в сообществе не обращаются в нуль и не возрастают до бесконечности, однако в нем не существует какого-либо устойчивого нетривиального решения. Тем не менее в таком сообществе ни один из видов не вымирает. Это означает, что модельные траектории ограничены сверху и снизу некоторыми положительными числами; однако, если рассматривать устойчивость в смысле Ляпунова, то сообщество неустойчиво, но, с другой стороны, оно устойчиво в смысле сохранения числа видов, т. е. в смысле второго экологического определения. Поэтому здесь мы будем говорить не об устойчивости сообщества, а об его экологической стабильности.
Остановимся несколько подробнее па требовании конечности численности. Поскольку в реальных сообществах неограниченного возрастания численности наблюдаться не может, то возникновение такого эффекта в модели говорит о ее неадекватности (не учтены какие-то лимитирующие факторы). Но адекватность мы обычно постулируем, и хотя неограниченное возрастание численности одного или нескольких видов вовсе не эквивалентно исключению видов из сообщества, но, тем не менее, этот эффект тоже можно рассматривать как нарушение его устойчивости. В пользу последнего говорит еще и следующее соображение: поскольку ресурсы, поступающие в экосистему, всегда ограничены, то неограниченное возрастание численности одного ИЗ видов приводит к тому, что ресурса будет не хватать для других, и эти виды с необходимостью ДОЛЖНЫ исчезнуть.
Еще по теме ВВЕДЕНИЕ:
- Введение в специальность.
- Введение
- Введение
- Введение
- Введение
- Введение
- Введение
- Введение в курс
- № 197-ФЗ, введенным в действие с 26 декабря 1995 г.
- № 197-ФЗ, введенным в действие с 26 декабря 1995 г.
- "Падение Запада" и глобальные проблемы человечества (общедоступное введение)
- Введение
- Введение
- Введение
- Введение
- Введение