Д. Последовательные рецепторы.
В некоторых особых случаях связывание агониста с его первичным рецептором само по себе еще не приводит к физиологическому ответу. Например, тиреоидный гормон L-тироксин (11.14) сначала связывается с рецептором на белке преальбумине.
Этот комплекс переносится кровью в щитовидную железу, где гормон взаимодействует с другим рецептором. По данным рентгеноструктурного анализа в молекуле преальбумина (ОММ 55 000) существуют два одинаковых канала, каждый из которых с высокой аффинностью (107 М-1) связывает по одной молекуле L-тироксина. При этом фенольная группа L-тироксина оказывается в середине белковой молекулы, а его аминокислотная часть — около входа в канал, так что его группа СОг~ сближается с группой NH3+ лизина. Средняя часть молекулы гормона (дифениловый эфир) жестко фиксируется в канале за счет тесного контакта с тремя «комплементарными карманами» в белке. В D-тироксине в отличие от L-изомера группа NH3+ располагается напротив группы NH3+ белка и кулоновское отталкивание ослабляет взаимодействие с рецептором [Andrea et al., 1980].Для стероидных гормонов конечным рецептором является ДНК, что следует из антагонистического действия актиномицина D (специфического ингибитора ДНК-зависимого синтеза РНК); их первые рецепторы — белки. Существование в матке, влагалище и передней доле гипофиза белков, специфичных для
эстрогенов, было показано в ранних работах Jensen, Jacobson. (1962) при изучении специфического связывания 17£1-[3Н]- эстрадиола с различными тканями. В фаллопиевых трубах существуют места избирательного связывания прогестерона.
Эти комплексы стероид — рецептор образуются в ядрах или проникают в них и связываются с хроматином (в отсутствие белка связывания стероидов не происходит). При этом депрес- сируется кодон ДНК, и на нем начинает синтезироваться мРНК.
В цитозоле рибосомы синтезируют по этой мРНК определенный белок. Все это происходит очень быстро, и физиологический ответ на гормон может проявиться уже через час. Известны и другие типичные примеры индукции синтеза характеристичных белков [Cohen, 1966].Основной андрогенный стероид, дигидротестостерон, избирательно связывается с кислыми белками в андрогензависимых тканях (например, в предстательной железе) [Anderson, Liao,. 1968]. В свою очередь кортикостероиды связываются со специфическими белками в цитоплазме клеток печени и других органов. Эти комплексы проникают в ядра, где связываются с ДНК, что приводит к синтезу специфических мРНК (их образование может быть подавлено актиномицином D), на которых и синтезируются в цитоплазме характеристичные ферменты [Sekeris, 1971]. По аналогичному механизму осуществляется диуретическое действие альдостерона [Edelman, Bogoroch, Porter, 1963].
2,3,7,8-Тетрахлордибензо-пара-диоксин (2.28а), широко известная примесь в трихлорфеноле, захватывается рецепторными белками в цитозоле печени и переносится в ядро, где он индуцирует цитохром Р-450 и арилгидроксилазу. Эти его свойства не являются вредными, но у некоторых линий мышей диоксин вызывает экспрессию генов, приводящую к дегенерации, делению и дифференциации клеток — так называемую «поддерживающую» патологию. Возможно, в этих процессах участвуют нормальные системы клеточной регуляции, но полной уверенности в этом нет. С аналогичными рецепторами взаимодействует большинство веществ, в молекулах которых имеется плоская поверхность площадью около 0,3X1 нм [Poland, Knutson, 1982]. Диоксин индуцирует также и б-левулинсинтетазу, фермент, определяющий скорость биосинтеза порфиринов. Степень токсичности диоксина для человека точно не установлена, возможно, что ее сильно преувеличивают. Однако учитывая его высокую токсичность для некоторых линий мышей, диоксин в настоящее время считают наиболее токсичным из всех низкомолекулярных соединений [Whitlock, Israel, 1964].
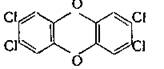
Диоксин
(2.28а)