4.0. Нуклеиновые кислоты
В 1944 г. Oswald, Avery и сотр. открыли, что дезоксирибонуклеиновая кислота (ДНК) пневмококков является носителем генетической информации, а ДНК одного штамма бактерий может передавать наследственную информацию другому (процесс трансформации) [Avery, MacLeod, McCarty, 1944].
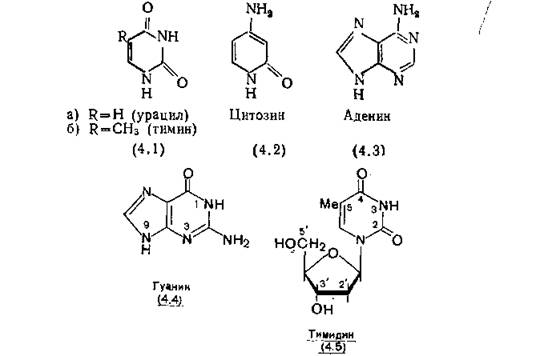
А. Дезоксирибонуклеиновая кислота (ДНК). Эта наиболее важная из нуклеиновых кислот встречается в митохондриях и хлорпластах, но большая часть ее содержится в ядре. ДНК является носителем всей генетической информации клетки. В зависимости от обстоятельств клетка использует определенную часть закодированной информации, хранящейся в ДНК в виде последовательности пиримидиновых-—тимин (4.1,6), цитозин (4.2) и пуриновых—аденин (4.3), гуанин (4.4) оснований. Цепь ДНК имеет остов, состоящий из остатков рибозы и фосфорной кислоты; к этому остову прикреплены азотистые основания. Молекула ДНК представляет собой двойную спираль, образованную двумя спирально закрученными цепями и скрепленную водородными связями между основаниями [Watson, Crick, 1953].
О значениях длины связей и величины их углов см. Wine и сото. (1980). ,
ОММ ДНК, полученной из различных источников, включая млекопитающих и бактерий, колеблется от 10 до 109 млн. Молекулы ДНК, содержащиеся в ядрах позвоночных, обычно образуют палочки около 3 мкм в длину и 0,18 нм в поперечнике. Неядерная ДНК (в митохондриях и бактериях), как правило, имеет циклическую структуру, причем у бактерий она одноцепочечная. Расстояние между плоскостями пиримидиновых и пуриновых оснований по вертикали около 0,33 нм (измеренные от центра до центра молекул), так что свободного пространства между ними нет [Jordan, 1968]. Пуриновые и пирамидиновые основания планарны, причем парные основания расположены в одной плоскости друг с другом и с С—Г и С—4' атомами сахара, с которыми они связаны, а плоскость рибозного цикла расположена почти под прямым углом к плоскости оснований.
Структура ДНК тесно связана с двумя ее главными функциями: репликацией генов (удвоение генов при синтезе ДНК) и транскрипцией (экспрессия генов при синтезе РНК) (рис. 4.1).
ДНК синтезируется в ядрах из мононуклеотидов ДНК-полимеразой. Этот процесс происходит на матрице, которой является другая молекула ДНК- Известны вещества, ингибирующие синтез ДНК благодаря своей способности связываться с матрицей, выводя ее таким образом из строя (например, акридины). Многие онковирусы, у которых РНК единственная нуклеиновая кислота, содержат обратную транскриптазу — полимеразу, синтезирующую ДНК на вирусной РНК [Temin, Mizutani, 1970]. В настоящее время ведутся поиски веществ, способных избирательно ингибировать этот фермент.
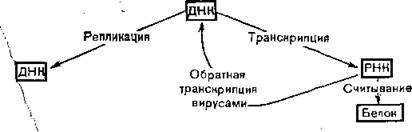
Рис. 4.1. Функции нуклеиновых кислот в живой клетке.
ДНК высших форм жизни в основном составляют перечисленные выше четыре основания. Единственное исключение 5-ме- тилцитозин — в ДНК растений он может заменять до 25% всего цитозина, однако в ДНК животных его значительно меньше, а в бактериальной ДНК всего 0—2% [Vanyushin et al., 1968]. Бактериальная и вирусная ДНК могут содержать и другие метилированные основания, такие как 6-метиладенин, 2-метиладе- нин или 5-гидроксиметилурацил. В некоторых фагах весь цитозин может быть заменен на 5-гидроксиметилцитозин, синтезируемый вирусиндуцируемым ферментом в бактерии-хозяине [Cohen, 1963].
В бактериальных ДНК при переходе от вида к виду может значительно изменяться соотношение оснований: отношение двух бифункциональных оснований (Г+Ц) к сумме двух монофункциональных (А + Т) может меняться в интервале 0,45—2,80. Следует отметить, что у высших растений и животных этот интервал 0,6—0,9 [Belozersky, Spirin, 1958].
Б. Хроматин. В ядре ДНК входит в состав длинных нитей плоских клинообразных нуклеосом. Каждая нуклеосома представляет собой двойную спираль из 150 пар оснований, в свою очередь закрученную вокруг 8 молекул основных белков (гистонов).
Очевидно, наружная часть нуклеосом доступна для РНК-полимеразы и кислых белков-репрессоров. В митозе эти нити нуклеосом сливаются в хромосомы. Приведенный пример относится к клеткам печени крысы и типичен для эукариот. Бактериальный хроматин имеет совершенно иную структуру [Finch et al., 1977].В. Рибонуклеиновые кислоты (РНК). ДНК не только выполняет функцию самовоспроизведения, но и служит матрицей для синтеза рибонуклеиновых кислот. Существуют три основных типа РНК: матричная (мРНК), транспортная (тРНК), иногда называемая растворимой РНК, и рибосомная (рРНК). Структуры ДНК и РНК очень схожи, но в РНК заспирализована только часть каждой молекулы и вместо дезоксирибозы в состав РНК входит рибоза, а вместо тимина — урацил. Все три типа РНК содержат также и метилированные основания. ОММ РНК всегда меньше, чем у исходной ДНК: у мРНК она обычно около 1 млн, а у тРНК — около 25000. Все типы РНК участвуют в процессе синтеза белков, различающегося у бактерий и высших организмов.
Матричные РНК — это фактически целое семейство РНК, и каждая из них содержит ту часть генетической информации, заложенной в ДНК, которая требуется клетке в данный момент ее жизнедеятельности (см. рис. 4.1). мРНК образует комплекс с рибосомами, где она определяет синтез специфичной для данного белка последовательности (см. ниже), после завершения которого комплекс мРНК с рибосомой распадается и сама мРНК разрушается. Выбор аминокислоты определяется /последовательностью расположения триплетов азотистых оснований в молекуле мРНК (кодоны), определяющей включение Данной аминокислоты в полипептидную цепь. Набор кодонов составляет генетический код, одинаковый для всего живого.
Существует целая группа транспортных РНК (тРНК), каждая из которых специфически этерифицирует только одну определенную аминокислоту. Эфиры аминокислот связываются с рибосомами в порядке, определяемом мРНК, проходящей в этот момент через рибосому (см. рис. 5.7). Рибосома способна, используя энергию гуанозинтрифосфата, присоединять каждую новую аминокислоту к концевой карбоксильной группе растущей полипептидной цепи.
Освободившаяся тРНК при этом возвращается в цитоплазму за другой молекулой аминокислоты.Более подробно этот процесс может быть представлен следующим образом: З'-гидроксильная группа рибозы (на концевом остатке аденозина молекулы тРНК) ферментативно ацилируется специфической к данной тРНК аминокислотой. Затем под контролем мРНК происходит амидирование этого эфира пеп- тидил-тРНК. После этого растущий пептид, связанный с тРНК на «донорном» участке рибосомы, оказывается связанным с аминоацильной тРНК на «акцепторном» участке. Это передвижение катализируется ферментом пептидилтрансферазой. Далее под действием транслокационного фермента происходит сдвиг растущей пептидной цепи вновь на донорный участок рибосомы. К освободившемуся акцепторному участку присоединяется следующая аминоацил-тРНК, и процесс повторяется. Концевой участок тРНК всегда представлен последовательностью ЦЦА, где Ц — остатки цитидиловой и А — адениловой кислоты.
Некоторые нуклеотиды тРНК содержат необычные основания. Например, аланиновая тРНК содержит гипоксантин, 1-ме- тилгипоксантин, 1-метилгуанин, 2-диметилгуанин, тимин (связанный с рибозой), дигидроурацил (три остатка) и два остатка псевдоурацила (ф), т. е. урацила, связанного с рибозой поэтому С—5, а не по атому азота. В тРНК обнаружены и другие минорные основания: 5-метилцитозин, 7-метилгуанин, 2-мстнл- гуанин и 1-метил-, 2-метил- и 6-метиладенины. В состав предшественников тРНК входят обычные пуриновые и пиримидиновые основания; под действием фермента метилазы, переносящей метальную группу от метионина, они метилируются in situ, в результате чего повышается специфичность тРНК. У млекопитающих метилирование азотистых оснований происходит только в ядрах и специфично для каждой ткани. В злокачественных опухбдях скорость метилирования резко возрастает, при этом образуются тРНК, не свойственные здоровой ткани [Kuchino, Borek, '[978].
Рибрсомная РНК (рРНК) связывается более чем с 50 раз- личными бейками в рибосомных субъединицах (разд.
5.4). Ее предшественник синтезируется в ядрышке и там же частично метилируемся. Более подробно о химии нуклеиновых кислот см. Davidson (1976) и Neidle (1982) *.Многие Эффективные противобактериальные, противопрото- зойные, противовирусные и противоопухолевые лекарственные вещества прямо или косвенно ингибируют синтез ДНК. Вполне попятно, что большинство из них вошло в клиническую практику до того, как стало известно их воздействие на генетический материал клетки — главный регулятор ее жизнедеятельности и наследственности. Строго направленное воздействие на вредную клетку очень долго оставалось недостижимой мечтой из-за существующей опасности повреждения генов полезной клетки. В настоящее время принято правило, разрешающее применение химического вещества в качестве лекарственного препарата, только если оно почти не влияет на синтез и устойчивость ДНК полезных клеток и не образует с ней ковалентных связей. Это предупреждает возможность мутаций в здоровых клетках при применении большинства существующих избирательных лекарственных веществ, которые обычно разделяют на пять классов.