§ 2, Вольтерровские модели и балансовые уравнения экологии
В современной литературе по математической экологии вольтерровскими моделями сообществ п видов принято считать системы
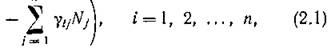
где ег — это скорость естественного прироста или смертности г'-го вида в отсутствие всех остальных видов, а знак и абсолютная величина (i ]) отражают соответственно характер и интенсивность влияния /-го вида на 1-й вид, уи — показатель внутривидового взаимодействия для г-го вида.
Матрицу Г = ||у/у||, отражающую структуру связей сообщества, часто называют матрицей сообщества-, с введенной в предыдущем параграфе знаковой матрицей S она связана соотношением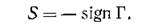
Система (2.1) была получена В. Вольтерра из следующих соображений: динамика г-го вида в отсутствие всех остальных видов описывается известным логистическим уравнением (как правило, уу > 0, что указывает на внутривидовую конкуренцию, или самолимитирование по численности і-го вида), а влияние /-го вида на i-й вид выражается членом, пропорциональным произведению N[Nj в соответствии с так называемой гипотезой «встреч и эквивалентов». Гипотеза «встреч и эквивалентов» полагает, что изменение численности і-го вида (жертвы) в результате поедания его /-м видом (хищником) определяется количеством встреч между особями данных видов, откуда и возникает произведение NiNj, а прирост численности хищника таков, как если бы осуществлялось немедленное преобразование съеденных особей жертвы в особи хищника с некоторым коэффициентом пропорциональности — «эквивалентом».
Гипотеза «встреч и эквивалентов», обязанная своим происхождением, по-видимому, принципу «встреч» в статистической физике, объясняет вид уравнений (2.1) в случае взаимодействий хищник — жертва и, очевидно, не способна обосновать его для других типов биологического взаимодействия.
Кроме того, в силу своей «небиологичности» эта гипотеза весьма уязвима для критики со стороны экологов. Тем не менее оказывается, что вольтерровские уравнения (2.1) могут быть получены и из более естественных предпосылок.Рассмотрим сообщество, структура которого изображена на рис. 21. Компоненты сообщества разобьем на три основные группы.
Продуценты с биомассами (или концентрациями) Х{  Это в основном зеленые растения, способные фиксировать световую энергию и использовать в пищу простые вещества.
Это в основном зеленые растения, способные фиксировать световую энергию и использовать в пищу простые вещества.
Консументы с концентрациями г/у К этой
группе отнесем животных, пожирающих другие организмы, а также разлагателей, расщепляющих мертвую органику на простые вещества, которые используются продуцентами.
Субстраты с концентрациями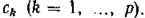 Это
Это
абиотические вещества (в основном продукты жизнедеятельности консументов), используемые продуцентами.
Составим уравнения, отражающие баланс масс каждой из этих компонент. По сути дела, эти уравнения есть не что иное, как законы сохранения массы. Эти балансовые экологические уравнения можно записать в таком виде: 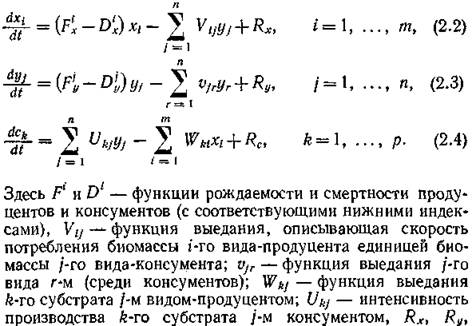
Rc — сумма входных и выходных потоков соответствующих компонент. Естественно, что все эти функции зависят и от параметров внешней среды (от сезонно изменяющейся влажности, температуры и т. п.).
Но такая модель страдает одним существенным недостатком — излишней общностью. Необходимо, исходя из различных биологических соображений, конкретизировать вид входящих в модель функций.
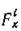
Будем считать, что рождаемость /-го вида зависит только от интенсивности светового потока I и от количества потребляемых субстратов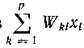 .
.
соображений, связанных с законами сохранения, можно считать, что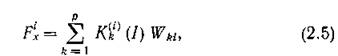
т. е. осуществляется стехиометрия с коэффициентами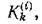 которые можно интерпретировать как долю fe-ro субстрата, идущего на воспроизводство биомассы /-го продуцента. Заметим, что если последовательно проводить стехиометрическую гипотезу, то
которые можно интерпретировать как долю fe-ro субстрата, идущего на воспроизводство биомассы /-го продуцента. Заметим, что если последовательно проводить стехиометрическую гипотезу, то
К = 1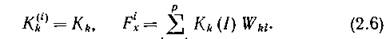
Для скорости прироста /-го консумента аналогично (2.5) получаем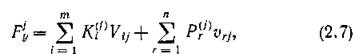
где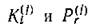 —стехиометрические коэффициенты взаимо
—стехиометрические коэффициенты взаимо
действия консументов с продуцентами и хищничества среди консументов.
Рассмотрим один частный вид этой общей модели. Пусть рождаемость продуцентов не лимитирована ни светом, ни минеральным питанием, а ограничена лишь чисто физиологическими пределами. Тогда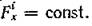 Поскольку компо
Поскольку компо
ненты-субстраты в этом случае не оказывают влияния на динамику остальных компонент, то в модели можно рассматривать только виды-продуценты и виды-консументы.
Аналогичная ситуация возникает и при лимитировании светом, поскольку свет является внешним фактором по отношению к этой системе, и при постоянстве светового потока Flx также можно считать постоянными. Рассматривая замкнутую систему, мы должны положить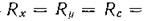
= 0.
Самое простое предположение о виде функций смертности — это гипотеза об их постоянстве.В реальных сообществах ситуация сложнее: как внутривидовая конкуренция, так и межвидовая конкуренция за один и тот же ресурс (пищу) приводит к увеличению смертности, когда численность соответствующих видов возрастает.
Описать этот механизм какими-либо балансовыми соотношениями навряд ли возможно. Поэтому самое простое предположение — это предположение о линейной зависимости функций смертности от численностей соответствующих видов:5 = 1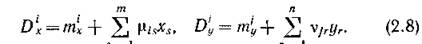 Г — I
Г — I
Здесь т‘л и пг’у — коэффициенты естественной (физиологической) смертности, цн и Vjj описывают интенсивность внутривидовой конкуренции; там, где конкуренция за общий ресурс отсутствует, соответствующие (1 И V должны быть равны нулю.
Какое предположение можно сделать о виде функций выедания? Самое простое — это гипотеза о линейной зависимости этих функций от концентрации (численности, биомассы) выедаемого вида
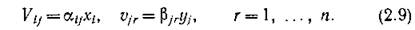
Конечно, предположение о линейной зависимости функций выедания от рациона справедливо в весьма узком диапазоне концентраций выедаемой пищи (например, для малых концентраций) и для такой трофической стратегии консумента, когда в рационе не делается какого-либо предпочтения одному виду перед другим. Но эти предположения, несмотря на их ограниченность, не противоречат биологии. Более того, они справедливы для весьма широкого класса экологических ситуаций. В общем случае, естественно, существуют как верхние физиологические границы количества потребляемой пищи (так что функции выедания должны
описываться зависимостями с насыщением), так и более сложные формы трофических стратегий (например, при трофическом предпочтении виды выедаются последовательно, а не одновременно; тогда сами функции выедания одного вида имеют более сложную структуру, зависящую от концентрации и других видов).
Учитывая сделанные предположения, модель (2.2)—(2.3) можно записать в виде
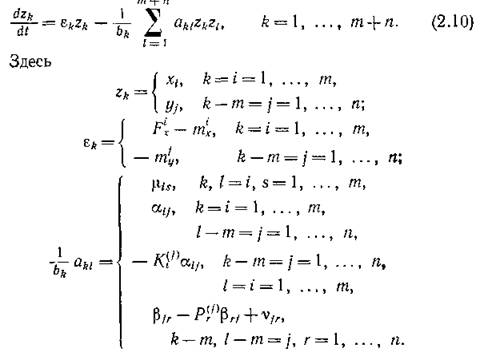
Система (2.10) (которая масштабным преобразованием переменных Nk = bkzk и введением матрицы с элементами ук1 = = ciki!(bkbi} может быть записана в форме (2.1)) представляет собой самый общий вид вольтерровских моделей.
В. Воль- терра подробно исследовал более частные случаи — с определенными ограничениями на матрицу ||й*,||. На примере системы «ге хищников — п жертв» мы постараемся выяснить, каков биологический смысл этих ограничений. Система имеет вид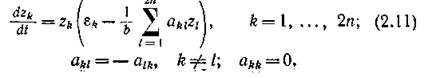
Пусть все хищники сосредоточены на одном, а все жертвы — на другом трофическом уровне, причем как внутривидовая, так и межвидовая конкуренция среди видов одного и того же уровня отсутствует. Тогда, если гх, .... z„ — численности жертв, — численности хищников, то
— численности хищников, то
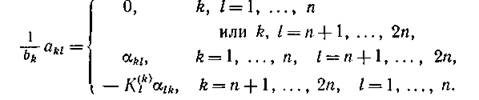
Отсюда следует, что ограничение типа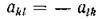 будет
будет
выполняться тогда и только тогда, когда
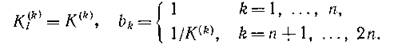
Что будет означать это условие? Обратимся снова к законам сохранения. Для этого рассмотрим ситуацию, когда т невозобновимых субстратов с концентрациями служат пищей для п видов с биомассами yj. Из законов сохранения следует, что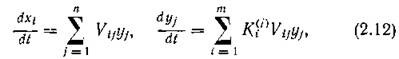
а суммарная масса субстратов и питающихся ими видов с учетом того, что только часть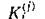 поглощенного
поглощенного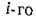 субстрата идет на воспроизводство /-го вида) должна быть постоянна. Это выполняется только тогда, когда для всех
субстрата идет на воспроизводство /-го вида) должна быть постоянна. Это выполняется только тогда, когда для всех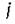 (строгая стехиометрия), и
(строгая стехиометрия), и
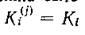
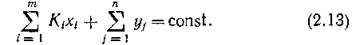
Такая форма записи означает, что только определенная часть массы каждого субстрата идет на образование новой биомассы потребляющего его вида (вне зависимости от того, какой это вид).
Все определяется энергетической и биохимической ценностью потребляемого субстрата, а видовая специфичность описывается выбором той или иной функции выедания.Возможен и иной подход. Пусть набор сбалансированного пищевого рациона определяется заданием функций выедания, а видовая специфичность задается тем, что различные виды тратят на воспроизводство своей биомассы разные доли своего рациона (/С(/)). Тогда закон сохранения запишется в виде
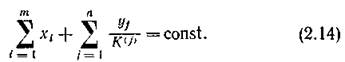
В вольтерровских моделях описывается именно такая ситуация. Трудно сказать, какой из этих двух подходов соответствует действительности. По-видимому, истина лежит где-то посредине. Для нас достаточно того факта, что вольтерров- ское описание биологически вполне осмысленно.