§ 3. Устойчивость в вольтерровских моделях сообщества
Под устойчивостью в моделях типа (2.10) понимается существование и устойчивость по Ляпунову положительного стационарного решения N* — наиболее жесткое из всех требований к «равновесной» траектории сообщества, которым можно придать экологический смысл.
Положительное стационарное решение системы (2.1) должно удовлетворять линейной алгебраической системе
где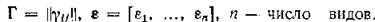 Пусть
Пусть
вектор е таков, что все компоненты системы (3.1) положительны, или, иными словами, N* принадлежит внутренности положительного ортанта Рп «-мерного пространства. Например, для системы (2.11) это означает, что во всяком случае 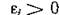 у всех жертв и ;
у всех жертв и ;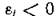 , у всех хищников.
, у всех хищников.
При исследовании систем типа (2.1) В. Вольтерра рассматривал два больших класса таких систем — консервативные и диссипативные. Система (2.1) называется консервативной (по В. Вольтерра), если существует такой набор положительных чисел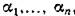 , что
, что 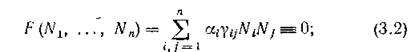
если же квадратичная форма F (Nlt ..., Nn) положительно определена, система называется диссипативной.
Биологический смысл этих определений состоит в следующем. Если величины а; отождествить со средними значениями биомассы особей каждого вида, то выражение
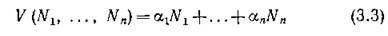
будет показывать общую биомассу сообщества.
Из уравнений (2.1) непосредственно вытекает, что скорость изменения общей биомассы есть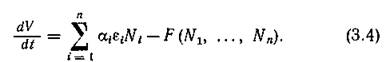
Первое слагаемое правой части (3.4) отражает влияние (постоянных в данном случае) причин прироста или убыли видов, а второе слагаемое — влияние взаимоотношений видов. Таким образом, консервативные сообщества характеризуются тем, что взаимодействие видов не влияет на изменение общей биомассы сообщества, а диссипативные системы — тем, что это взаимодействие замедляет прирост общей биомассы.
Поскольку для консервативной системы, очевидно, выполняются соотношения
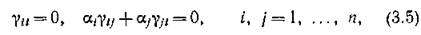
т. е.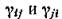 противоположны по знаку, то ясно, что консер
противоположны по знаку, то ясно, что консер
вативными могут быть лишь сообщества, единственным ‘ типом межвидовых отношений в которых (не считая нейтрализма) выступают отношения хищник — жертва, а само- лимитирование видов отсутствует. В. Вольтерра показал, что необходимым и достаточным условием консервативности системы (2.1) является следующая совокупность требований к элементам матрицы Г: 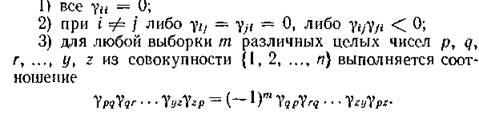
Таким образом, за исключением случая п = 2, консервативность требует выполнения некоторых соотношений типа равенств между параметрами системы, что делает консервативность маловероятным свойством для реальных систем. Консервативные сообщества в математической экологии — это такая же идеализация, как и системы без трения в механике, и аналогично тому, как нередко в механике пренебрегают трением, для некоторых сообществ правомерна приближенная гипотеза об их консервативности.
Основное свойство консервативных систем (2.1)—это существование первого интеграла на траекториях, лежащих в положительном ортанте, в случае невырожденности матрицы Г (при четном ft):
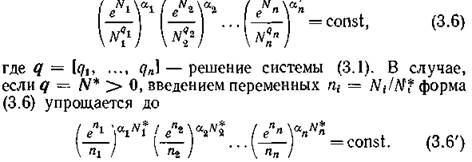
Используя эти соотношения, В.
Вольтерра показал, в частности, что, когда q > 0, все виды ограничены сверху и снизу положительными константами и, если начальное состояние системы отлично от N*, по крайней мере один вид имеет незатухающие колебания численности. При этом асимптотические средние величинт. е.
отлично от N*, по крайней мере один вид имеет незатухающие колебания численности. При этом асимптотические средние величинт. е. выражения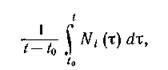
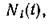
имеют при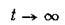 пределы, совпадающие с их стационар
пределы, совпадающие с их стационар
ными значениями (закон асимптотических средних).
Поведение траекторий консервативной системы (2.1) можно интерпретировать с позиций первого метода Ляпунова анализа устойчивости равновесия системы N*. Действительно, легко видеть, что функция 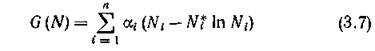
является функцией Ляпунова для точки N* в области минимальное значение , , достигается в точке N* и про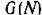 изводная вдоль тоаекторий системы
изводная вдоль тоаекторий системы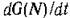 равна нулю
равна нулю
всюду в области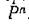 . Таким образом, равновесие N* устойчиво по Ляпунову (но не асимптотически).
. Таким образом, равновесие N* устойчиво по Ляпунову (но не асимптотически).
Второй метод Ляпунова, основанный на линеаризации системы в точке равновесия и анализе спектра полученной матрицы, позволяет исследовать характер траекторий в малой окрестности равновесия.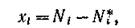 Полагая
Полагая
получаем линейную систему
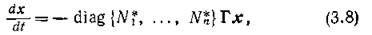
а если рассматривать отклонения в форме
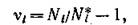
то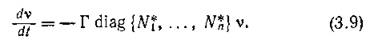
Поскольку линейная замена переменных не изменяет спектра матрицы линейной системы, матрицы систем (3.8) и (3.9) имеют одинаковый спектр.
Можно показать, что при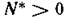 этот спектр состоит из чисто мнимых чисел.
этот спектр состоит из чисто мнимых чисел. Действительно, если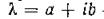 — собственное значение
— собственное значение
матрицыdiag{...)r с собственным вектором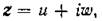
то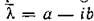 также является собственным значением с
также является собственным значением с
собственным вектором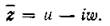 Тогда, по определению
Тогда, по определению
собственного вектора,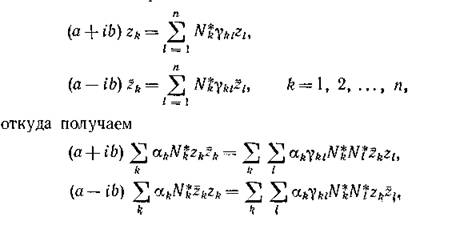
Сложив почленно эти равенства, после соответствующей перегруппировки слагаемых правой части будем иметь
К. К I
в силу (3.5). Отсюда, так каг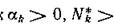 0 и не все zk = О,
0 и не все zk = О,
необходимо следует, что а = 0, что и требовалось [13]).
Таким образом, линейное приближение траекторий в окрестности равновесия — это суперпозиция п/Ъ синусоид с различными, вообще говоря, периодами. Здесь мы имеем типичный пример, иллюстрирующий возможности обоих методов Ляпунова в случае чистой мнимости спектра: прямая линеаризация не дает определенного ответа об устойчивости или неустойчивости N*, которая может быть установлена лишь анализом соответствующей функции Ляпунова.
В случае, когда не все компоненты qi решения системы стационарного состояния положительны, можно утверждать, что один или несколько видов исчезают или их численность растет неограниченно. В противном случае, т. е. если все численности ограничены сверху и снизу положительными константами, существуют положительные асимптотические средние для всех Nh которые, как можно показать, всегда равны соответствующим корням (/; системы (3.1).
Полученное противоречие доказывает утверждение.Таким образом, существование положительного равновесия выступает не только достаточным, но и необходимым условием стабильного функционирования консервативного сообщества.
Обратимся теперь к диссипативным сообществам. Из определения диссипативной системы следует необходимое условие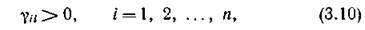
т. е. все виды должны обладать самолимитированием. Если вспомнить, что у консервативных систем все уи = 0, становится ясным, что определения консервативной и диссипативной системы несовместимы. Ясно также, что существуют
системы, не являющиеся ни консервативными, ни диссипативными— например, это системы, в которых лишь часть видов обладает самолимитированием.
Следующее необходимое условие диссипативности состоит в том, что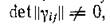 откуда вытекает единствен
откуда вытекает единствен
ность решения системы уравнений стационарного состояния. И наконец, все главные (т. е. симметричныеотносительно главной диагонали) миноры матрицы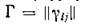 должны
должны
быть положительны.
Для диссипативной системы (2.1) В. Вольтерра аналогично (3.6) получил соотношение

[1] То же верно и для матрицы Г, что дает еще одно необходимое усдовие диссипативности системы (2,1),
локально асимптотически устойчиво, и характер сходимости возмущенных траекторий к N* известным образом зависит от набора собственных чисел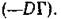
Заметим, что на траекториях системы (2.1) для функции 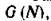 введенной в (3.7), справедливо равенство
введенной в (3.7), справедливо равенство
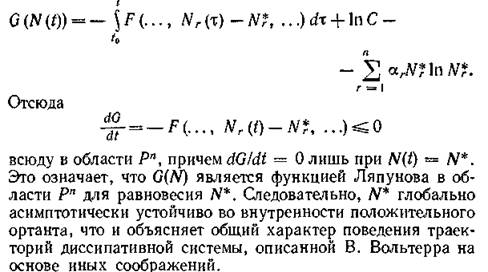
Что касается случая, когда не все qr положительны, то если отрицательный корень только один, соответствующий вид исчезает *); если таких корней несколько, то среди соответствующих видов по крайней мере один должен исчезать. Таким образом, и для диссипативных систем так же, как и для консервативных, существование устойчивого равновесия N* > 0 служит не только достаточным, но и необходимым условием стабильного функционирования сообщества.