§ 4. Структуры хищник — жертва
Как отмечалось в предыдущем параграфе, консервативными могут быть лишь сообщества, состоящие из видов хищник — жертва без самолимитирования. Из условий (3.5) вытекает, что симметричные элементы матрицы Г должны иметь противоположные знаки; в частности, Г может быть антисимметричной.
Для устойчивости консерва
тивного сообщества необходимо и достаточно, чтобы система уравнений стационарного состояния (3.1) с невырожденной матрицей Г имела положительное решение. Ясно, что выбором подходящих значений et- этого всегда можно добиться; например, достаточно положить е = Г[1, 1, ..., 1И. Таким образом, устойчивость консервативного сообщества сводится лишь к существованию положительного решения (3.1), а если по условиям задачи допустимы вариации е, то — к невырожденности матрицы сообщества Г. При этом оказывается, что устойчивость может быть установлена с помощью анализа только трофического графа сообщества. Результаты настоящего параграфа показывают, что сообщество хищник — жертва (без самолимитирования) устойчиво лишь тогда, когда множество видов допускает такое разбиение на непересекающиеся пары, что в каждой паре один из видов питается другим.
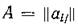
Всякой действительной п х «-матрице можно
поставить в соответствие ориентированный граф с п вершинами так, что вершины іи / связаны ребром / -> г тогда и только тогда, когда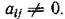 Для матриц Г консерватив
Для матриц Г консерватив
ных сообществ очевидно, что если есть ребро / —> і, то существует и ребро с противоположным направлением, поэтому структуру сообщества можно изобразить неориентированным графом.
Такой граф будем обозначать G(T).Будем говорить, что некоторое подмножество Р множества всех ребер G(T) задает разбиение на пары хищник — жертва, если ни одна из вершин, принадлежащих ребрам Р, не служит вершиной более чем одного ребра. Разбиение на пары хищник — жертва назовем полным разбиением, если в нем участвуют все п вершин графа. Графы, изображенные на рис. 22, а и б, не имеют полного разбиения; во втором из них добавление ребра 2—3 приводит к возникновению полного разбиения (рис. 22, б); в графе рис. 22, г оно может быть выполнено тремя различными способами.
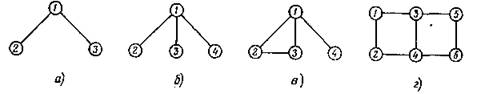
Рис. 22. Графы сообществ хищник—жертва.
Теорема 1. Пусть А — антисимметричная матрица, G(A) — (неориентированный) граф, соответствующий А. Тогда если А — невырожденная, то в G(4) существует полное разбиение (на пары хищник. — жертва). Обратно, если в G(A) есть полное разбиение, то существует невырожденная антисимметричная матрица В такая, что
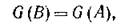
и элементы В могут быть выбраны сколь угодно близкими к соответствующим элементам А.
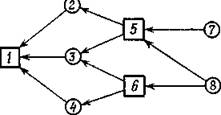
Рис. 23. Граф с трофическими уровнями. Стрелки идут от жертвы к хищнику.
Эта теорема, доказательство которой будет приведено ниже, показывает, что наличие полного разбиения является необходимым условием устойчивости сообщества с антисимметричной матрицей А. Это условие и достаточно в том смысле, что если А — вырожденная, то сколь угодно близкая к А матрица В уже невырожденная и дает устойчивое сообщество. Теорема 1 позволяет сразу утверждать, что сообщество с графом заметил еще В. Вольтерра, устойчивость таких систем возможна лишь при четных п); неустойчива и система рис. 22, б.
рис. 22, а неустойчиво (как
В структуре многих реальных сообществ хищник — жертва, как правило, выделяются трофические уровни — совокупности видов, питающихся только видами следующего уровня и служащие пищей видам предыдущего уровня.
Граф рис. 23 дает пример такого сообщества, где вид 1 питается на видах 2, 3 и 4, пищей которым служат виды 5 и 6 и т. д. В случае, если в графе наличие трофических уровней не очевидно, полезным оказывается следующее определение.Будем называть графб(Л) трофическим, если существует такое подмножество видов Т, что ни один из них не питается видами Т, а виды, не принадлежащие Т, питаются только на видах Т. Для многоуровневых систем это формальное определение, очевидно, выполняется, так как в качестве Т можно взять объединение уровней, находящихся на нечетном «расстоянии» от жертв самого последнего уровня (на рис. 23 вершины Т обозначены квадратиками).
Теорема 2. Если А — невырожденная матрица и граф G(A) — трофический, то в G(A) существует полное оазбиение.
Последнее условие не выполнено для графа рис. 23, так что в нем не существует полного разбиения и, следовательно, соответствующая антисимметричная матрица вырождена.
Обратимся теперь к доказательству сформулированных теорем. Пусть А — антисимметричная матрица и полное разбиение б(Л) задается подмножеством ребер Р. Определим матрицу В (г) следующим образом:

Тогда 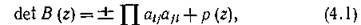
где произведение берется по всем парам элементов А, соответствующим ребрам Р, и полином р(г) таков, что р(0) = 0. Тогда det В(0) 0 и вообще det B(z) =£ 0, за исключением
конечного множества (не более чем п) значений г. Положим г0 достаточно близким к 1 (не равным 1, если det В(1) = 0). Тогда искомая матрица есть
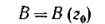
и вторая часть теоремы 1 доказана.
Заметим, что доказательство не требует антисимметричности А, а использует лишь свойство
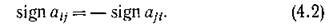
Пусть теперь det Л ^о.
По стандартному правилу матрице А поставим в соответствие ориентированный граф G'(A). В разложении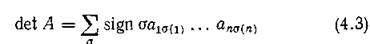
найдется такая перестановка а чисел от 1 до п, что соответствующий член определителя
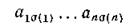
отличен от нуля. Этой перестановке отвечает подграф графа G', содержащий все п вершин, каждая цз которых является началом одного ребра и концом только одного ребра в Go. Тогда по теореме Эйлера [14])
графа G', содержащий все п вершин, каждая цз которых является началом одного ребра и концом только одного ребра в Go. Тогда по теореме Эйлера [14])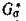 представляет собой объединение не связанных между собой ориентированных циклов. Заметим, что для каждого ребра в Ga в силу свойства (4.2) найдется противоположно направленное ребро в
представляет собой объединение не связанных между собой ориентированных циклов. Заметим, что для каждого ребра в Ga в силу свойства (4.2) найдется противоположно направленное ребро в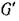 и, следовательно, неориентированное ребро межпу теми же вершинами в G(A). Теперь, если все циклы имеют четные длины, то полное разбиение G(A) на пары хищник — жертва задаетсямножеством Р тех ребер, которые соответствуют ребрам
и, следовательно, неориентированное ребро межпу теми же вершинами в G(A). Теперь, если все циклы имеют четные длины, то полное разбиение G(A) на пары хищник — жертва задаетсямножеством Р тех ребер, которые соответствуют ребрам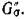
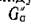
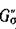
Если граф G(A) — трофический, то все циклы должны иметь четную длину (каждый цикл представляетсобой некоторое число переходов из множества вершин ТъТ и обратно), и теорема 2 доказана.
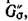
Допустим теперь, что для всякой перестановки ст, дающей ненулевой член в разложении (4.3), в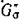 найдется цикл нечетной длины.
найдется цикл нечетной длины.
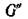 с циклами по тем же вершинам, что и циклы
с циклами по тем же вершинам, что и циклы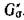 , а суммиро-
, а суммиро- вание членов (4.3) по этому классу дает определитель некоторой антисимметричной матрицы Лс с элементами
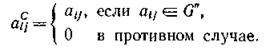
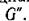
Переупорядочив виды должным образом, получим Ас в блочно-диягональном виде с блоками, соответствующими циклам Согласно предположению среди них найдутся циклы нечетной длины и, поскольку антисимметричный определитель нечетного порядка равен нулю, определитель Ас, равный произведению определителей блоков, также равен нулю. Тем самым 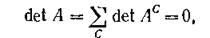
и полученное противоречие завершает доказательство теоремы 1.
При большом числе видов в структуре сообщества хищник — жертва поиск полного разбиения может оказаться затруднительным. Положение облегчается тем, что существуют специальные алгоритмы, которые позволяют установить разбиение на пары, захватывающие максимальное число вершин графа, а следовательно, и полное разбиение, если оно существует.