§ 5. Типы трофических функций хищника и устойчивость системы хищник — жертва
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что а'х < 0 и а (х) = О при X < оо, то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции чгаенностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь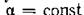 (так что, в отличие от предыдущей
(так что, в отличие от предыдущей
модели,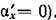 Естественно, что решения (2.1) зависят
Естественно, что решения (2.1) зависят
от конкретного вида трофической функции V (%), которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
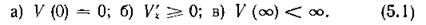
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
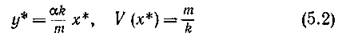
при естественном ограничении mlk < V (оо).
Существует еще одна стационарная точка (0, 0), соот
ветствующая тривиальному равновесию.
Нетрудно показать, что эта точка — седло, а сепаратрисами являются координатные оси.Характеристическое уравнение для точки (%*, у*) имеет вид
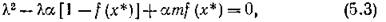
где
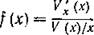
Очевидно, что для классической вольтерровской модели 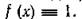 Поэтому величину f можно рассматривать как
Поэтому величину f можно рассматривать как
меру отклонения рассматриваемой модели от вольтер- ровской.
При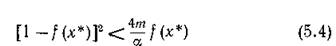
стационарная точка (%*, у*) — фокус, и в системе появляются колебания; при выполнении противоположного неравенства — узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
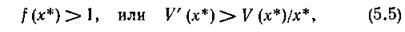
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник — жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией V(x)/x, условие возрастания которой (положительность производной) выглядит как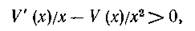
или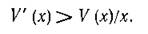
Последнее условие, взятое в точке х*, есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности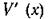 оно должно выполняться и в некотопой окрестности точки
оно должно выполняться и в некотопой окрестности точки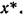 Таким образом, если — чис
Таким образом, если — чис
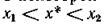
ленности жертв в этой окрестности, то
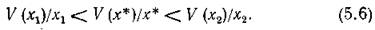
Пусть теперь трофическая функция V имеет вид, изображенный на рис.
11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений х (поскольку V (х) выпукла вверх)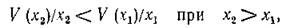
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической 1
функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности і
как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант — возникновение устойчивого предельного цикла — невозможен, что легко доказывается. (
В самом деле, выражение j
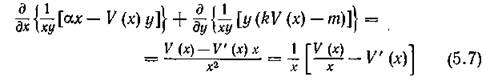 I
I
в положительном квадранте всегда положительно, если только V (%) имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака [10]), в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла і
существовать не может. (
Итак, можно сделать вывод: если трофическая функция I
имеет вид, изображенный на рис. 11, а, то хищник не может I
быть регулятором, обеспечивающим стабильность популя- і
ции жертвы и тем самым устойчивость всей системы в целом.
Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые.
С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник — жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов-насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.
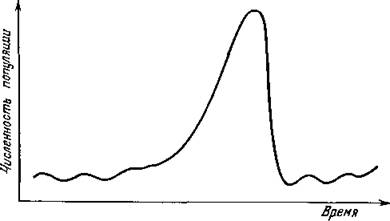
Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.—Austr. J. Zool., 1964, 12, № 3,
p. 362—380.)
Если же в систему «хищник — жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, б. Точка хп на этом графике — точка касания прямой, проведенной из начала координат, графика трофической функции V (х). Очевидно, что в этой точке функция W (х) = V (х) !х имеет максимум. Также легко показывается, что для всех х* < х„ выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв х* меньше хп, будет асимпто
тически устойчиво. Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник — жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении х (при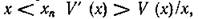 при
при
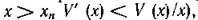 то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.
то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.