§ 2. Разомкнутые и замкнутые трофические цепи
До сих пор мы не конкретизировали понятие «ресурс» — в реальных экосистемах это может быть либо энергия, либо какие-то жизненно необходимые вещества (например, углерод, азот или фосфор).
Если по отношению к энергии любая экосистема является незамкнутой, энергия солнечного света «протекает» через экосистему, попутно рассеиваясь в виде тепла (расходы на дыхание, на метаболическую активность и т. д.), то по отношению к биологическим веществам (углероду, азоту, фосфору и т. д.) экосистемы являются замкнутыми (с той или иной степенью замкнутости). Замыкание происходит за счет деятельности так называемых «разла- гателей» (микроорганизмы, грибы, черви), которые разлагают мертвую органику до минеральных компонентов, служащих питательными веществами для первичных трофических уровней. То же самое справедливо и для трофических цепей. Поэтому в дальнейшем мы будем рассматривать два типа трофических цепей: незамкнутые («проточные») и замкнутые («циклы»). Схематически оба эти типа изображены на рис. 28.Рост и развитие экосистем, как правило, лимитируется каким-либо фактором (принцип Либиха). Это может быть либо солнечный свет, либо какой-нибудь химический элемент — обычно азот или фосфор. Если солнечный свет — ресурс невозобновимый, то химические вещества за счет деятельности разлагателей снова вовлекаются в круговорот. Поэтому в том случае, когда лимитирующим фактором оказывается солнечный свет, трофическая цепь незамкнута, а если это один из биогенных элементов, то трофическая цепь замыкается в цикл.
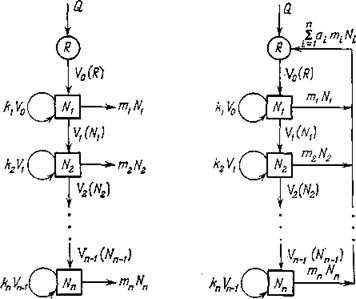
Рис. 28. Схема трофической цепи длины п: а) незамкнутая («проточная») цепь; б) замкнутая цепь («цикл»). Коэффициенты а,- (О йй 1) — доли восстановленного вида- ми-разлагателями ресурса, содержащегося в отмершей биомассе ('-го вида. Эти коэффициенты характеризуют степень замыкания трофической цепи.
Рассмотрим более подробно схемы на рис. 28. Здесь R — ресурс, используемый 1-м видом с биомассой . Удельная скорость использования I — это количество ресурса потребляемое единицей биомассы (одной особью) 1-го вида
. Удельная скорость использования I — это количество ресурса потребляемое единицей биомассы (одной особью) 1-го вида
за единицувремени. Из общего количества потребляемого ресурса лишь к^я доля его идет на воспроизводство
лишь к^я доля его идет на воспроизводство
новой биомассы 1-го вида, остальное расходуется на поддержание жизнедеятельности. Кроме того, с постоянной скоростью т1 биомасса 1-го вида отмирает, 2-й вид использует уже в качестве ресурса биомассу 1-го вида, потребляя ее с удельной скоростью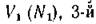 вид — биомассу вто
вид — биомассу вто
рого и т. д. Цепочка заканчивается на л-м виде, биомассу которого уже никто не потребляет.
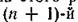
Вторая схемя этого писунка отличается от первой тем, что существует вид — разлагатель, который в ка
честве ресурса использует мертвую биомассу первых п видов и за счет своей жизнедеятельности частично восполняет убыль ресурса /?. Мы будем предполагать, что разложение мертвой биомассы происходит достаточно быстро и высвободившийся при разложении ресурс практически мгновенно снова может быть использован 1-м видом. Кроме того, скорость роста биомассы вида-разлагателя столь велика, что он также практически мгновенно может утилизировать любое количество мертвой биомассы других видов. В этом случае нет необходимости рассматривать численность или биомассу вида-разлагателя в качестве отдельной фазовой переменной — можно считать, что вся отмирающая биомасса (или некоторая ее часть — если учитывать неизбежные потери в этом процессе) сразу же возвращается в блок ресурса.
Пусть экосистема имеет трофический граф типа изображенных на рис. 28. Предположим, что вся таким образом организованная экосистема стремится к некоторому состоянию равновесия, причем в этом состоянии отличны от нуля стационарные численности только первых q видов. Такое устойчивое равновесие естественно называть трофической цепью длины q.
Пусть скорость поступления в экосистему внешнего ресурса постоянна и равна Q. Ставится следующий вопрос: какова должна быть эта скорость (при заданных трофическихфункциях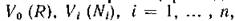 и параметрах
и параметрах
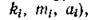 чтобы в таком сообществе существовало устойчивое нетривиальное равновесное состояние с ненулевыми численностями первых q видов? Другими словами, каковы условия существования трофической цепи длины
чтобы в таком сообществе существовало устойчивое нетривиальное равновесное состояние с ненулевыми численностями первых q видов? Другими словами, каковы условия существования трофической цепи длины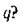 Очевидно, эти условия определяются параметрами экосистемы
Очевидно, эти условия определяются параметрами экосистемы
и скоростью поступления в экосистему внешнего ресурса.
По трофическим графам на рис. 28 достаточно просто выписываются следующие балансовые уравнения для биомасс видов.
а) Незамкнутая цепь:
і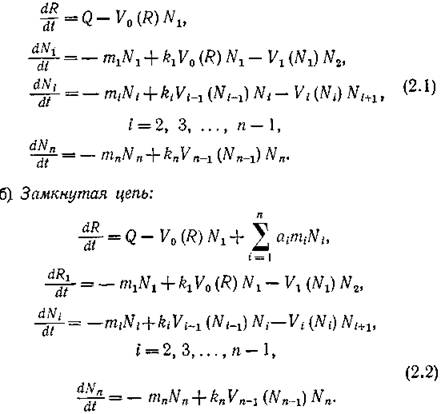
По своему биологическому смыслу параметрыи удовлетворяют ограничениям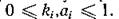
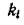
Если считать, что ни один вид не имеет в избытке трофического ресурса, т. е. трофические связи «напряжены» (см. стр. 95), то в этом случае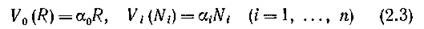
и уравнения (2.1) и (2.2) переходят в уравнения вольтер- ровского типа (за исключением первых уравнений, которые содержат член Q). Тогда, полагая формально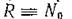 и
и
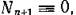 мы получим для описания динамики трофических цепей двух типов следующие две системы,
мы получим для описания динамики трофических цепей двух типов следующие две системы,
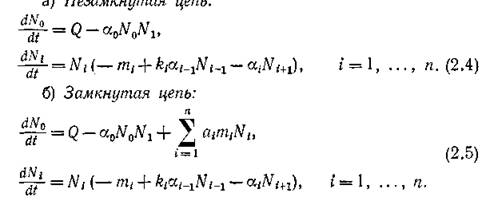
В последующих параграфах будут рассмотрены условия существования и устойчивости равновесий в этих системах, или, иными словами, условия существования трофических цепей.