§ 2. Классическая модель Вольтерра
Пусть X (/) И у (0 — численность жертв И ХИЩНИКОВ соответственно. Предположим, что единственным лимитирующим фактором, ограничивающим размножение жертв, является давление на них со стороны хищников, а размножение хищников ограничивается количеством добытой ими пищи (количеством жертв).
Тогда в отсутствие хищников численность жертв должна расти экспоненциально с относительной скоростью а, а хищники в отсутствие жертв — также экспоненциально вымирать с относительной скоростью т. Коэффициенты а и т — коэффициенты естественного прироста жертв и естественной смертности хищников соответственно.Пусть V = V (х) — количество (или биомасса) жертв, потребляемых одним хищником за единицу времени, причем k-я часть полученной с этой биомассой энергии расходуется хищником на воспроизводство, а остальное тратится на поддержание основного обмена и охотничей активности. Тогда уравнение системы хищник — жертва можно записать в виде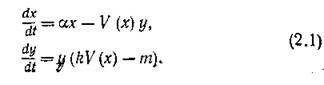
Функцию V (х) обычно называют трофической функцией хищника или функциональным откликом (functional res- ропсе) хищника на плотность популяции жертвы. Именно эти функции обычно определяются в экспериментальных работах, посвященных изучению хищничества, и к настоящему времени считается установленным, что эти функции обычно принадлежат к одному из следующих трех типов (рис. 11). По-видимому, динамическое поведение системы в значительной степени зависит от вида трофической функции.
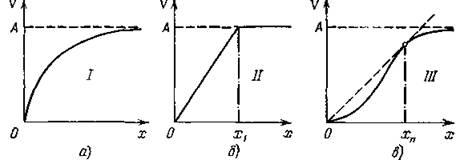
Рис. 11. Различные типы трофических функций в системе хищник — жертва: а) этот тип характерен для беспозвоночных и некоторых видов хищных рыб; б) трофическая функция с резко выраженным порогом насыщения характерна для хищников-фильтраторов (например, многих моллюсков); в) такой тип характерен для позвоночных — организмов, проявляющих достаточно сложное поведение (например, способных к обучению).
Аналогичный вид будет иметь трофическая функция, если жертвы могут вырабатывать защитную стратегию (например, прятаться в убежище, недоступное хищникам).При малых, значениях х, например, когда трофические отношения в системе напряжены и почти все жертвы становятся добычей хищника, который всегда голоден и насыщения не наступает (ситуация довольно обычная в природе), трофическую функцию V (х) можно считать линейной функцией численности жертв, т. е. V = (Зх. Кроме того, предположим, что k = const. Тогда
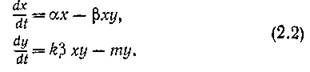
Система (2.2) с точностью до обозначений совпадает с классической моделью хищник — жертва В. Вольтерра,
который показал, что эта система имеет интеграл вида

В системе (2.2) имеется еще одно положение равновесия — начало координат. Нетрудно видеть, что эта точка — седло. Оси кооординат являются сепаратрисами, причем ось Оу входит в седло, а ось Ох — выходит из него (см. рис. 12).
Несмотря на то, что модель Вольтерра смогла объяснить многие реально наблюдавшиеся явления, у нее есть большой недостаток — негрубость (в математическом смысле этого слова) вольтерровских циклов, так что при любых сколь угодно слабых возмущениях фазовых координат система переходит с одного цикла на другой. По-видимому, более адекватные модели должны обладать этим свойством «грубости».
С точки зрения теории устойчивости состояние равновесия системы (х*, у*) — это состояние безразличного равновесия, устойчивое по Ляпунову, но не асимптотически. Отсутствие асимптотической устойчивости равновесия указывает на то, что в вольтерровской системе отсутствуют механизмы, стремящиеся сохранить ее нетривиальное равновесное состояние.
Высказанное выше утверждение об устойчивости равновесия достаточно легко доказывается построением соответствующей функции Ляпунова. Но об этом в следующем параграфе.