§ 4. Глобальные экстремальные свойства траекторий сообщества с горизонтальной структурой
Если сформулированный в предыдущем параграфе принцип носит локальный характер (говорить о монотонном возрастании мощности можно, строго говоря, только в достаточно малой окрестности положения равновесия), то принцип, который мы сформулируем в этом параграфе, будет справедлив глобально, т.
е. для любой траектории сообщества, принадлежащей положительному ортанту. Доказывать его проще для модели в численностях, т. е. для классической формы вольтерровских уравнений


Из этого выражения видно, что по мере приближения к стационарному состоянию скорость движения все более и
[1] См., например, к а р р Ч., X о у в Ч. Количественные методу принятия решений в управлении и экономике. Перевод с англ. — М.: Мир, 1966, с. 296.
[1] См., например, Барбашин Е. А, Введение в теорию устойчивости. — М.: Наука, 1967, с. 46—47.
более замедляется, хотя вдали от этого состояния она может быть достаточно велика.
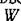
Как связаны между собой функции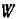 и веденная в предыдущем параграфе F? Вычисляя значение.. в устойчивой стационарной точке и учитывая, что
и веденная в предыдущем параграфе F? Вычисляя значение.. в устойчивой стационарной точке и учитывая, что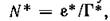 получим
получим
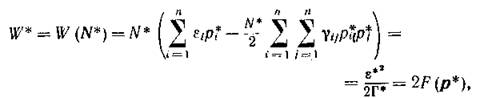
т. е. максимальные значения этих двух функций пропорциональны.
Все эти результаты не только подтверждают данное ранее качественное описание поведения диссипативных вольтерровских систем, но и попускают содержательную
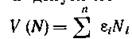
интерпретацию. Величина характеризует,
< = і
по сути дела, скорость прироста биомассы в случае, когда конкуренция и какое-либо лимитирование по ресурсу отсутствуют, и этот прирост определяется только физиологическими репродуктивными возможностями организмов и их естественной смертностью. Поэтому естественно назвать V репродуктивным потенциалом сообщества. Аналогом этого понятия в экологической литературе может служить понятие общей продукции экосистемы. Выражение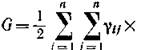
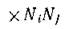
можно рассматривать как меру скорости рассеяния энергии в результате межвидовой и внутривидовой конкуренции, т. е. суммарного расхода биомассы сообщества на конкуренцию. Поэтому мы будем называть величину G общими затратами сообщества на конкуренцию, или, опять же возвращаясь к чисто экологическим формулировкам, затратами на дыхание экосистемы. Поэтому возрастание W в процессе эволюции сообщества можно интерпретировать так, что сообщество стремится максимизировать разность между своим репродуктивным потенциалом и общими затратами на конкуренцию. Это можно сделать несколькими путями: максимизируя репродуктивный потенциал при заданных затратах на конкуренцию либо
минимизируя затраты на конкуренцию при ограниченном репродуктивном потенциале (существуют и некоторые промежуточные ситуации). Как показывают наблюдения, в природе реализуются все эти случаи. Например, так называемые r-конкуренция и К-конкурещия, которым соответствуют два противоположных механизма регулирования численности.
Происхождение этих терминов становится ясным, если мы обратимся к несколько иной форме записи уравнений (4.1) (см.
§ 2 гл. VI):
естественного прироста і-говида, то о видах, выигрывающих конкурентное соревнование за счет увеличения собственной плодовитости, говорят, что они используют г-стратегию. А поскольку Кі — это емкость среды для і-го вида, то о видах, выигрывающих конкурентное состязание за счет оптимального использования ресурсов (использование широкого спектра ресурсов, уменьшение затрат на конкуренцию благодаря «разумному» его распределению и т. и.), говорят, что они используют К-стратегию. Таким образом, можно сказать, что r-стратегия сообщества заключается в максимизации репродуктивного потенциала, а К-стратегия — в минимизации общих затрат на конкуренцию.
И в заключение дадим еще одну интерпретацию доказанного в предыдущем параграфе утверждения о том, что функция F(p) в равновесии достигает максимума. Пусть процесс эволюции сообщества квазистационарен по общей численности, т. е. при медленной эволюции композиции сравнительно быстро устанавливается квазиравновесие по
численности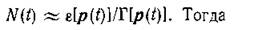
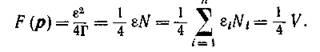
Отсюда сразу следует, что сообщество эволюционирует к состоянию с такой композицией, при которой его репродуктивный потенциал максимален. Однако этот принцип является локальным.