§ 2. Формализация понятия ниши и динамические уравнения конкуренции
В экологической литературе существует несколько не совпадающих определений экологической ниши. С точки зрения математика, наиболее подходящим из них следует считать такое, которое, отражая существо дела, допускает возможность формализации.
Удачным компромиссом такого рода является определение ниши, опирающееся на такие Понятия, как спектр ресурса и функция потребления.Допустим, что ресурс, потребляемый биологическими видами, характеризуется неким (в общем случае многомерным) параметром х (размер потребляемой пищи, видовой ее состав, пространственные координаты местообитания и т. д.), а количество доступного для потребления ресурса с характеристиками х определяется некоторой функцией К (х). Тогда множество значений х с заданной на нем функцией К (х) называется спектром (или пространством) ресурса.
Пусть, кроме того, потребление ресурса данным видом характеризуется некоторым вероятностным распределением плотности f (х) — функцией потребления — со средним значением х0 и конечной дисперсией о2. Тогда экологическая ниша вида определяется точкой х0 на спектре ресурса и заданной функцией f (х) — плотностью случайного распределения вокруг точки х0. Например, особи какого-то вида охотнее всего потребляют пищу размера х0, потребление пищи иного размера подчиняется некоторому — теоретическому или эмпирическому — вероятностному закону с плотностью f (х). Предпочитаемый размер х0 естественно считать центром ниши, а среднеквадратичное отклонение а — шириной ниши, характеризующей степень специализации данного вида по потреблению ресурса х: малые значения а— узко специализированный вид, большие а — широко специализированный.
Ясно, что, увеличивая размерность вектора х, можно добиться теоретически полной адекватности введенного определения условиям существования биологического вида.
Однако, ради наглядности и упрощения количественного анализа моделей, дальнейшие рассмотрения будут предполагать одномерность спектра ресурса. Пути обобщения полученных результатов в большинстве случаев естественны и очевидны.Когда мы рассматриваем сообщество нескольких видов, конкурирующих за общие ресурсы, естественно считать — и это подтверждается наблюдениями, — что отвечающие разным видам точки ха удалены друг от друга на какие-то конечные расстояния. При этом само явление конкуренции, связанное с перекрыванием экологических ниш конкурирующих видов, в рамках принятого формализма возникает как результат пересечения областей определения соответствующих функций потребления fi (%) в пространстве
ресурса. Простейший (идеализированный) пример такого рода изображен на рис. 36, где ft (%) представляют собой одинаковые по форме кривые нормального распределения, расположенные равномерно вдоль спектра ресурса с расстоянием d между средними значениями; /С (х) = const. Если w — общая для всех кривых величина среднеквадратичного отклонения, то отношение wld можно рассматривать как меру близости экологических ниш, или меру плотности видовой упаковки сообщества, и по условиям устойчивости судить о допустимых пределах перекрывания ниш.
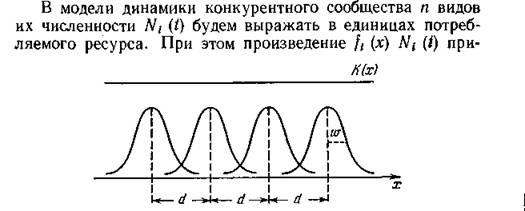
Рис. 36. Одномерный спектр ресурса К(х) и перекрывание экологических ииш, заданных нормальными функциями потребления.
обретает смысл той доли всего ресурса, потребляемого і-м видом, которая берется в точке х спектра ресурса. Тогда
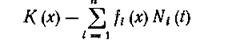
есть не что иное, как локальная (на спектре ресурса) разница между потенциально доступным и необходимым для сосуществования [//і (t), ..., N„ (01 количеством ресурса. Пока ресурс не лимитирует прирост численностей видов (при малых численностях), можно считать, что популяция увеличивается в соответствии с собственным коэффициентом естественного прироста г{ согласно мальтузианскому закону
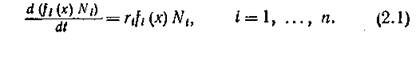
Лимитирующее влияние конкуренции можно выразить относительной мерой истощения ресурса в точке х:
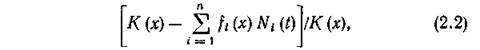
после чего динамика 1-го вида в точке х его экологической ниши описывается уравнением
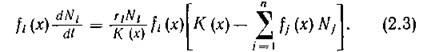
Интегрирование (2.3) по всему спектру ресурса дает
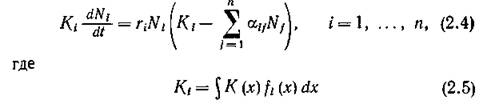
имеет смысл общего количества ресурса, потребленного 1-м видом — так называемая емкость ниши, а коэффициент конкуренции между t-м и /-м видами
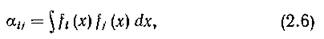
который пропорционален полной (по всему спектру) вероятности столкновения потребительских интересов данной пары видов в одной и той же точке спектра, характеризует степень перекрывания их экологических ниш.
Деля обе части (2.4) на Д;, получаем систему уравнений

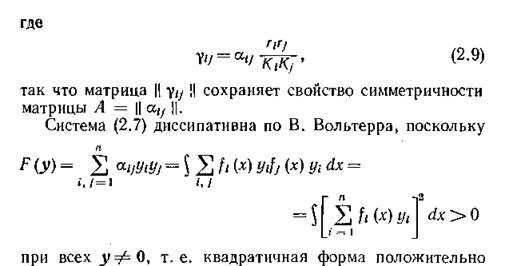
определена. Из диссипативности вытекает, что если в (2.7) существует положительное стационарное решение, то оно асимптотически устойчиво всюду в положительном ортанте Рп, и, более того, устойчивость этого равновесия является не только достаточным, но и необходимым условием стабильного функционирования сообщества п конкурирующих видов. Таким образом, критерием стабильности конкурентного сообщества являются условия существования положительного равновесия, т. е. положительного решения линейной системы
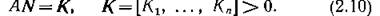
Ниже, в § 4 мы дадим «геометрическую» интерпретацию положительности решения (2.10), а пока рассмотрим совокупность конкурирующих видов с гауссовскими функциями пптпрЛлримя ГТ.ПЯ кяжппгп инпя


печивая формально асимптотическую устойчивость равновесия, но стремится к нулю быстрее любой конечной степени d/w, имея, таким образом, существенную сингулярность в нуле. Это порождает крайне длительное время возвращения возмущенной траектории к равновесному состоянию.
Кроме того, прямые расчеты на ЭВМ, выполненные для различных п в пределах п = 100, показывают, что при
малых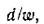 т. е. при значениях а, достаточно близких к 1, среди компонент решения (2.10) появляются уже отрицательные числа (когда все правые части одинаковы). Это значит, что с увеличением плотности видовой упаковки для сохранения стабильности необходимо и специальное изменение функции К (х) — такое, чтобы набор емкостей ниш сохранял положительность решения (2.10). Подробнее мы остановимся на этом в следующих параграфах.
т. е. при значениях а, достаточно близких к 1, среди компонент решения (2.10) появляются уже отрицательные числа (когда все правые части одинаковы). Это значит, что с увеличением плотности видовой упаковки для сохранения стабильности необходимо и специальное изменение функции К (х) — такое, чтобы набор емкостей ниш сохранял положительность решения (2.10). Подробнее мы остановимся на этом в следующих параграфах.